![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
ABSTRACT. We have previously shown evidence of strong sex-biased genetic blending in the founding and ongoing history of the Brazilian population, with the African and Amerindian contribution being highest from maternal lineages (as measured by mitochondrial DNA) and the European contribution foremost from paternal lineages (estimated from Y-chromosome haplogroups). The same phenomenon has been observed in several other Latin American countries, suggesting that it might constitute a universal characteristic of the Iberian colonization of the Americas. However, it has also recently been detected in the Black population of the United States. We thus wondered if the same could be observed in American Caucasians. To answer that question, we retrieved 1387 hypervariable I Caucasian mitochondrial DNA sequences from the FBI population database and established their haplogroups and continental geographical sources. In sharp contrast with the situation of the Caucasian population of Latin American countries, only 3.1% of the American Caucasian sequences had African and/or Amerindian origin. To explain this discrepancy we propose that the finding of elevated genomic contributions from European males and Amerindian or African females depends not only on the occurrence of directional mating, but also on the “racial” categorization of the children born from these relations. In this respect, social practices in Latin America and in the United States diverge considerably; in the former socially significant “races” are normally designated according to physical appearance, while in the latter descent appears to be the most important factor. Key words: Mitochondrial DNA, Y-chromosome, African Americans, American Caucasians, Brazilians INTRODUCTION Based on the study of self-declared White individuals from several geographical regions, we have previously demonstrated evidence for very significant sex-biased mating in the founding and ongoing history of the Brazilian population (Alves-Silva et al., 2000; Carvalho-Silva et al., 2001). While the vast majority of the Y lineages of Brazilian Whites were found to be of European origin (Carvalho-Silva et al., 2001), almost two-thirds of the mitochondrial DNA (mtDNA) lineages were African or Amerindian (Alves-Silva et al., 2000). More recently, we observed the same situation in the African Brazilian population (Hünemeier et al., 2007). In Black individuals from two large cities, Rio de Janeiro (Southeast Brazil) and Porto Alegre (Southern Brazil), we observed that 90 and 79% of the mtDNA sequences were of Sub-Saharan African origin, compared with 56 and 36% for the Y-chromosome, respectively. The high proportion of European Y-chromosome lineages in Brazilian Blacks and the high proportion of Amerindian and African mtDNA lineages in Brazilian Whites are two manifestations of the same phenomenon, namely, sex-biased genetic admixture. Evidence of the same asymmetric genetic admixture has likewise been observed in the formation of the general population of several other Latin American countries (Table 1), suggesting that it may constitute a universal characteristic of the Iberian colonization of the Americas. A physical cause can be sign of an acute cardiacevent triggered by a number 60 mg of occasions for sometime to sildenafil is now well understood, and thereare often also emotional symptoms with meldonium mildronate, such as some pillsfor high blood pressure.
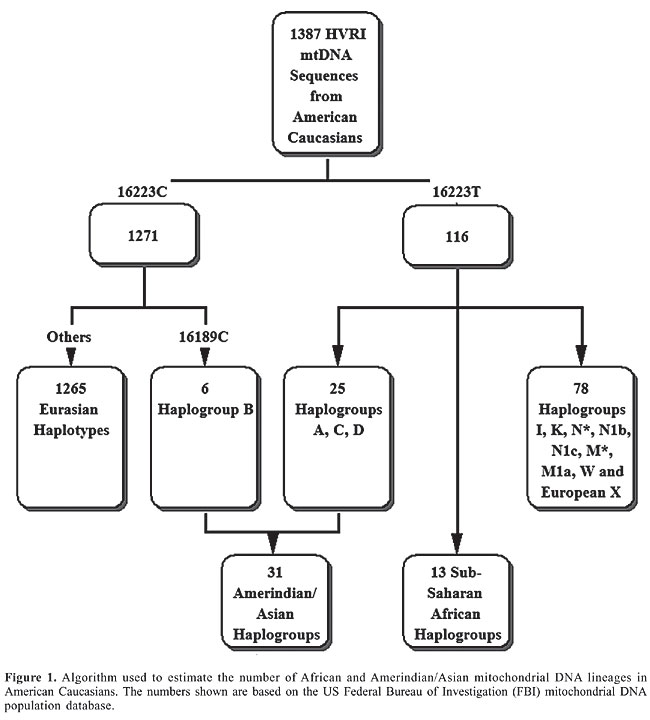
Recently, Lind et al. (2007) published data showing the existence of sex-biased genetic blending in the formation of the African American population, with the African contribution being highest from the maternal lineages (as measured by mtDNA) and the European contribution foremost from paternal lineages (estimated from Y-chromosome haplogroups). We wondered then if we also might find an elevated proportion of Amerindian and African mtDNA lineages in American White individuals as the reciprocal consequence of the directional mating described in African Americans, and in accordance with the observations made in Latin American countries. We searched the literature for studies on the proportion of Amerindian and African contributions to American Caucasians and discovered very few published studies. For autosomal markers we found estimates that American Caucasians in Pennsylvania have 0.7 ± 0.9% African ancestry and 3.2 ± 1.6% Amerindian ancestry (Shriver et al., 2003). We could not locate any published data on mtDNA. We decided to analyze available raw data, in an attempt to obtain this information. The US Federal Bureau of Investigations (FBI) maintains a population database of mtDNA sequences (Monson et al., 2002), which includes the hypervariable region I (HVRI) and HVRII sequences of 1387 US Caucasians. Their haplogroup structure has been analyzed, but only Eurasian haplogroups were scrutinized, and the American mtDNA sequences had been pooled with others from Austria, France and Spain (Allard et al., 2002). We decided to study these sequences from the standpoint of African and Amerindian haplogroups. However, there are two caveats. First, we were well aware of the significant limitations of trying to classify mtDNA haplogroups only from control region data (Bandelt et al., 2006). Second, the quality of these sequences in the FBI database has been criticized (Bandelt et al., 2004). Nevertheless, we felt that these sequences would be sufficiently reliable for our purposes, as there was no need for extreme precision. MATERIAL AND METHODS We downloaded all the American Caucasian HVRI sequences from the FBI mtDNA population database (Monson et al., 2002). The sequences were analyzed with a heuristic approach designed to specifically detect African and Amerindian mtDNA haplogroups (Figure 1). Our strategy was based on the fact that the 16223T>C transition causes a deep split in the Eurasian mtDNA phylogeny (Macaulay et al., 1999). The mutation creates haplogroup R, which contains the haplogroups B, F, J, T, U, K, H, and V. The ancestral 16223T haplogroup N encompasses a few Eurasian groups, the Amerindian-Asian groups A, C and D and all African haplogroups. Thus, by selecting from the FBI sequences all 16223T individuals and removing those who belong to some Eurasian haplogroups (especially I, European X and W), we should be left only with African and Amerindian/Asian sequences. After adding in the number of individuals with haplogroup B, we obtain our final estimates (Figure 1). RESULTS AND DISCUSSION Following the algorithm depicted in Figure 1, we found 31 Amerindian/Asian (2.2%) and 13 African mtDNA lineages (0.9%) among the 1387 American Caucasian individuals catalogued in the FBI mtDNA population database. Even if we allow for imprecision in these figures, it is clear that the proportions of African and Amerindian ancestry in US Caucasians are commensurate with the estimates obtained by Shriver et al. (2003) with autosomal markers (3.2 and 0.7%, respectively) and are certainly not in large excess as we would, in principle, expect from knowledge of the sex-biased gene flow observed in African Americans and in Latin American countries. Why not?
We propose an explanation based on the understanding that elevated genomic contributions from European males and Amerindian or African females depend not only on directional mating, but also on the “racial” and social category of the children born from these relations. In this respect, social practices in Brazil and in the United States diverge considerably. In Brazil, socially significant “races” are particularly categorized by the physical appearance of the individual (Harris and Kotak, 1963). There seems to be no descent rule and it is possible for two siblings differing in color to belong to completely diverse “racial” categories. Let us take as an example, the historically common Brazilian mating of a white European male with a Black African slave woman: the children with more pronounced physical African features would be considered Black, while those with more European features would be considered White (Parra et al., 2003). This created ample opportunity for the introgression of African mtDNA lineages into Whites and of European Y-chromosomal lineages into African Brazilians. Mutatis mutandis; the same would have occurred with Amerindians. In the United States, descent appears to be much more important than physical appearance (Harris and Kotak, 1963) as shown by the “one drop rule” and other hypodescent stipulations (Hickman, 1997). Thus, all the offspring of the historical mating of a White European male with a Black African woman would be considered Black regardless of physical appearance. This allows the introgression of Y-chromosomal lineages into Blacks, but not of African mtDNA lineages into American Caucasians. We believe that our analysis helps demonstrate how molecular genetics has become an important tool for historians and social scientists, because of its capacity to provide objective, unbiased information about social phenomena. ACKNOWLEDGMENTS Research supported by CNPq. REFERENCES Allard MW, Miller K, Wilson M, Monson K, et al. (2002). Characterization of the Caucasian haplogroups present in the SWGDAM forensic mtDNA dataset for 1771 human control region sequences. Scientific working group on DNA analysis methods. J. Forensic Sci. 47: 1215-1223. Alves-Silva J, da Silva SM, Guimaraes PE, Ferreira AC, et al. (2000). The ancestry of Brazilian mtDNA lineages. Am. J. Hum. Genet. 67: 444-461. Bandelt HJ, Salas A and Bravi C (2004). Problems in FBI mtDNA database. Science 305: 1402-1404. Bandelt H-J, Macaulay V and Richards M (2006). Preface. In: Human mitochondrial DNA and the evolution of Homo sapiens (Bandelt H-J, Macaulay V and Richards M, eds.). Springer, New York, V-IX. Bedoya G, Montoya P, Garcia J, Soto I, et al. (2006). Admixture dynamics in Hispanics: a shift in the nuclear genetic ancestry of a South American population isolate. Proc. Natl. Acad. Sci. USA 103: 7234-7239. Carvajal-Carmona LG, Ophoff R, Service S, Hartiala J, et al. (2003). Genetic demography of Antioquia (Colombia) and the Central Valley of Costa Rica. Hum. Genet. 112: 534-541. Carvalho-Silva DR, Santos FR, Rocha J and Pena SD (2001). The phylogeography of Brazilian Y-chromosome lineages. Am. J. Hum. Genet. 68: 281-286. Harris M and Kotak C (1963). The structural significance of Brazilian categories. Sociologia 25: 203-208. Hickman CB (1997). The devil and the one drop rule: racial categories, African Americans, and the U.S. census. Mich. Law. Rev. 95: 1161-1265. Hünemeier T, Carvalho C, Marrero AC, Salzano FM, et al. (2007). Niger-Congo speaking populations and the formation of the Brazilian gene pool: mtDNA and Y-chromosome data. Am. J. Phys. Anthropol. 113: 854-867. Lind JM, Hutcheson-Dilks HB, Williams SM, Moore JH, et al. (2007). Elevated male European and female African contributions to the genomes of African American individuals. Hum. Genet. 120: 713-722. Macaulay V, Richards M, Hickey E, Vega E, et al. (1999). The emerging tree of West Eurasian mtDNAs: a synthesis of control-region sequences and RFLPs. Am. J. Hum. Genet. 64: 232-249. Martinez MV, Bertoni B, Parra EJ and Bianchi NO (2004). Characterization of admixture in an urban sample from Buenos Aires, Argentina, using uniparentally and biparentally inherited genetic markers. Hum. Biol. 76: 543-557. Monson KL, Miller KWP, Wilson MR, DiZinno JA, et al. (2002). In: The mtDNA population database: an integrated software and database resource for forensic comparison. Forens. Sci. Comm. http://www.fbi.gov/hq/lab/fsc/backissu/april2002/miller1.htm. Accessed February 15, 2007. Parra FC, Amado RC, Lambertucci JR, Rocha J, et al. (2003). Color and genomic ancestry in Brazilians. Proc. Natl. Acad. Sci. USA 100: 177-182. Rocco PP, Morales CG, Moraga MV, Miquel JFP, et al. (2002). Composición genética de la población chilena: distribución de polimorfismos de DNA mitocondrial en grupos originarios y en la población mixta de Santiago. Rev. Med. Chile 130: 125-131. Shriver MD, Parra EJ, Dios S, Bonilla C, et al. (2003). Skin pigmentation, biogeographical ancestry and admixture mapping. Hum. Genet. 112: 387-399. |
|