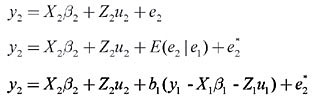![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
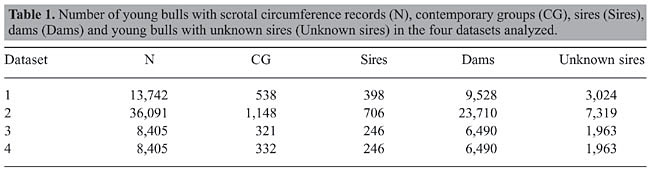
ABSTRACT. Data of pregnancy diagnosis from 24,945 Nellore heifers, raised under tropical conditions in Brazil and exposed to breeding at about 14 months of age, were analyzed simultaneously with 13,742 (analysis 1), 36,091 (analysis 2), 8,405 (analysis 3), and 8,405 (analysis 4) scrotal circumference (SC) records of contemporary young bulls in order to estimate heritability (h2) for yearling heifer pregnancy (HP) and for SC measured at around 15 (SC15) and 18 (SC18) months of age and to estimate genetic correlation between HP and SC15 (SC18). Heifer pregnancy was considered as a categorical trait, with the value 1 (success) assigned to heifers that were detected as pregnant by rectal palpation approximately 60 days after the end of a 90-day breeding season and the value 0 (failure) otherwise. In analyses 1 and 3, SC was measured at around 15 months of age and in analysis 2 and 4 it was measured at around 18 months of age. Only 8,848 animals from datasets 1 and 2 were common in both files, which means the same animals measured at different ages. Datasets used in analyses 3 and 4 included the same animals, measured at 15 and at 18 months of age, respectively. Heritability estimates for HP were similar in all analyses, with values ranging from 0.66 ± 0.08 to 0.67 ± 0.008. For SC15, the estimates were 0.57 ± 0.05 in analysis 1 and 0.60 ± 0.07 in analysis 3. For SC18, the estimates were 0.53 ± 0.03 in analysis 2 and 0.64 ± 0.06 in analysis 4. The estimates of genetic correlation between HP and SC15 were 0.15 ± 0.10 in analysis 1 and 0.11 ± 0.11 in analysis 3. For the correlation between HP and SC18, the values were 0.27 ± 0.10 in analysis 2 and 0.16 ± 0.11 in analysis 4. Based on standard errors and confidence intervals, the best heritability and genetic correlation estimates were obtained from analysis 2, which included more data and a better pedigree structure. Pearson correlation between HP and SC breeding values was similar to the genetic correlation estimates obtained from two-trait models, when all animals in the pedigree file were considered for its calculation. If only sires were considered for the calculation, Pearson correlation was higher but the pattern was the same as from two-trait analyses. The high heritability estimates obtained in the present study confirm that expected progeny difference (EPD) for HP can be used to select bulls for the production of precocious daughters and that the low genetic correlation between SC and HP indicates a greater efficacy of selection based on heifer pregnancy EPD than of selection based on scrotal circumference EPD. The results of the present study, although not conclusive, indicate that SC measured at around SC18 would have a higher genetic correlation with HP than would SC measured at around SC15. Key words: Expected progeny difference, Genetic correlation, Heritability, Method Â, Non-linear model, Sexual precocity INTRODUCTION In beef cattle, reproductive traits are a major concern, especially in harsh tropical environments. Nogueira (2004) presented a review on male and female reproduction traits of Zebu cattle (Bos indicus), especially in South American production systems. From this review and also as reported by Martin et al. (1992), Zebu heifers historically reach puberty at an older age than heifers of Bos taurus breeds. Age at puberty in extensive management systems is usually estimated from the age at first calving (AFC). However, Pereira et al. (2002) reported that AFC in Nellore cattle was to a great extent dependent on the age at which the heifers are exposed to breeding. In the Brazilian beef production systems, heifers are usually exposed to breeding at two years old but, according to Fajersson et al. (1991), Zebu heifers can reach puberty at yearling and can calve at 27 months of age if fed according to NRC protein recommendations. Doyle et al. (1996) and Evans et al. (1999) defined heifer pregnancy (HP) as the probability of a heifer being pregnant at the end of the breeding season when she was exposed to a bull or inseminated. Eler et al. (2002) reported a high heritability (0.57 ± 0.11) for HP in Nellore heifers exposed to breeding at about 14 months of age, and Silva et al. (2005) also reported high heritability for the trait in Nellore heifers exposed to breeding at about 16 months of age. The estimates reported by these authors were 0.52 ± 0.08 and 0.45 ± 0.03, in analyses using method  and a Bayesian approach, respectively. Although the results above indicate the use of HP expected progeny difference (EPD) to select for sexual precocity, it is still seldom used in the Brazilian production system. On the other hand, the majority of farmers pay attention to scrotal circumference (SC) measurements when buying young bulls. In Brazil, there is no unique national genetic evaluation for the Nellore breed. Besides the Brazilian Zebu Breeders Association (ABCZ), private groups (e.g., CFM, 2005; Lôbo et al., 2005; PAINT, 2005) have their own genetic evaluation programs and publish “Sire Summaries”. SC EPD is included in their publication. It is well known from the literature that SC can be used as an indicator of precocity. It is an easy and inexpensive trait to measure, has high heritability (Eler et al., 1996; Quirino and Bergmann, 1998) and is reported to be favorably associated with age at puberty (Brinks et al., 1978; Smith et al., 1989) and with age at first calving (Martins Filho and Lôbo, 1991; Pereira et al., 2000; Silva et al., 2005). SC is, however, no more than an indicator of age at puberty and the magnitude of its relationship with sexual precocity is yet to be well established. Evans et al. (1999), analyzing a small amount of data on Hereford cattle, reported an estimate close to zero for the genetic correlation between SC and HP. In Nellore cattle, Eler et al. (2004) used a two-trait animal model to analyze a large amount of data of SC and HP simultaneously. In addition to confirming a high heritability for HP (from 0.63 ± 0.08 to 0.69 ± 0.09), they reported a genetic correlation of 0.20 between the two traits. In Eler’s analysis, SC was measured from 400 to 650 days of age and adjusted to 450 days. The results from the literature, noted above (high heritability for HP and for SC, positive relationship between SC and AFC, and positive genetic correlation between SC and HP), lead to the conclusion that sexual precocity is a heritable trait and may respond to selection. There is, therefore, a need for finding good selection criteria for such traits. No conclusive study was found in the literature, relating the age of measurement of SC with the correlation between this trait and HP (or age at puberty) in Nellore or other Bos indicus cattle. Therefore, the main objective of the present study was to analyze a dataset of HP simultaneously with SC records taken at around 15 months of age and also the same dataset simultaneously with SC records taken at around 18 months of age to determine the genetic correlation between the two traits in Bos indicus cattle raised under tropical conditions in Brazil. MATERIAL AND METHODS Data The data were obtained from three herds owned by Agro-Pecuária CFM Ltda. CFM owns close to 17,000 Nellore cows and sells about 2,000 young bulls per year of 7,000 males weaned. Two herds were located in northeastern part of São Paulo State (20° 17’ 62” S and 49° 51’ 23” W; 20° 34’ 03” S and 50° 13’ 49” W), and the third one was located in the western part of Mato Grosso do Sul (20° 17’ 36” S and 55° 26’ 22” W), Brazil. Cattle management Both bulls and heifers were maintained on high-quality pasture (composed approximately of 40% Brachiaria brizantha, 50% Panicum maximum and 10% other grasses) and supplemented with salt and mineral (11% Ca, 6% P, 1% Mg, 4% S, 16% Na, 0.15% Cu, 0.15% Mn, 0.45% Zn, 0.015% I, 0.007% Co, and 0.002% Se). Calves were born from late August to December and remained with their dams up to seven months of age in a high-quality pasture. The 90-day heifer breeding season started in November and ended in January. Using multiple sire or single-sire breeding pastures, heifers were assigned by birth date to breeding management groups with heifers of similar age. The ratio of heifers per bull was about 35:1. All heifers were evaluated for pregnancy by rectal palpation, approximately 60 days after the end of the breeding season. At the start of the breeding season, all heifers in the herd were exposed to bulls regardless of their weight or body condition. Heifers that failed to conceive at 14 months were retained until the next breeding season when they were exposed as 2-year-old heifers. After new pregnancy diagnosis, they were removed from the herd at that time if they failed to conceive. Cows older than 2 years that did not conceive or cows with poor progeny performance were culled. Bulls were selected based on an index including standardized EPD for weaning weight, post-weaning gain, scrotal circumference, and muscle score, weighted 20, 40, 20, and 20%, respectively. Attention was paid to age at first calving and birth weight EPD. Since 2000, scrotal circumference EPD was replaced by heifer pregnancy EPD in the index for selection purposes, but not to rank young bulls for selling. Heifer pregnancy data HP was defined as the observation that a heifer conceives and remains pregnant upon palpation, given that she was exposed to breeding. Pregnant heifers or heifers with a calving record were given a score of 1 and non-pregnant heifers a score of 0. The data set contained 24,945 records of heifers born from 1993 to 2003 and exposed to breeding at about 14 months of age from 1994 through 2004. The heifers were daughters of 413 sires and 16,117 dams. Due to the use of multi-sire pastures, sires were not known for 6,901 heifers from those groups. Of the 24,945 records, 3,836 (15.4%) were scored 1, and 21,109 (84.6%) were scored 0. Scrotal circumference data Four datasets were obtained for scrotal circumference: Dataset 1 included records from 13,742 young bulls born from 1986 through 2003 in the same herds as heifers. SC was measured at about 15 months of age (SC15). Dataset 2 included records from 36,091 young bulls born from 1984 through 2003 in the same herds as heifers. SC was measured at about 18 months of age (SC18). Only 8,848 young bulls from dataset 1 and 2 were present in both files. Datasets 3 and 4 included, besides records from datasets 1 and 2, 8,405 young bulls born from 1997 through 2003 and measured at 15 and 18 months, respectively. Due to the use of multi-sire pastures, some young bulls came from those groups and had unknown sires. Descriptive statistics for the datasets are presented in Table 1.
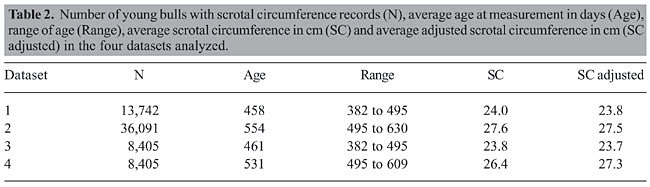
Statistical analysis Records from datasets 1 and 3 were adjusted to 450 days and those from datasets 2 and 4 were adjusted to 550 days using a segmented polynomial technique (Gallant and Fuller, 1973). This procedure gave a better adjustment curve in a previous study (Mourão et al., 2004). The age of SC measurement and average SC are presented in Table 2.
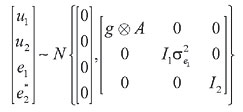
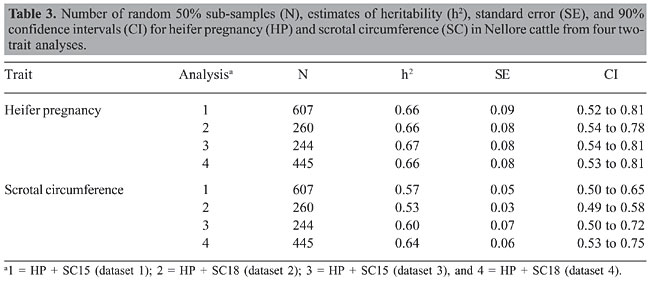
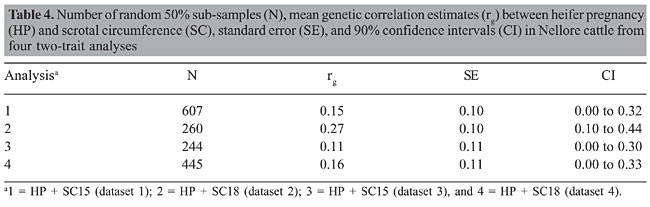
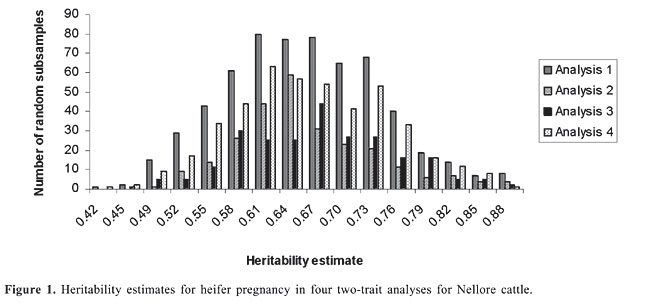
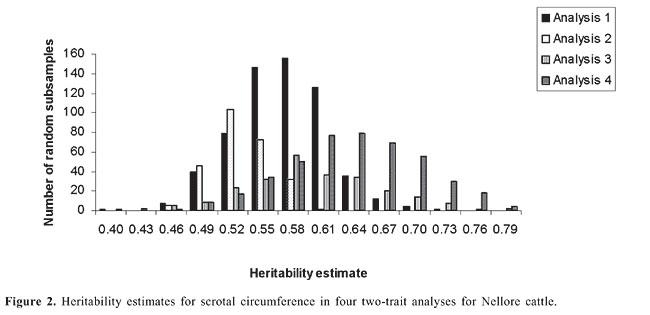
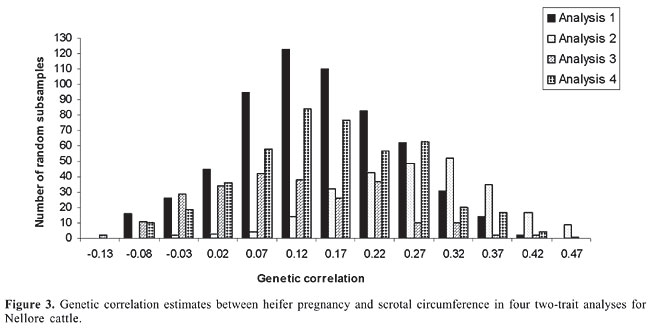
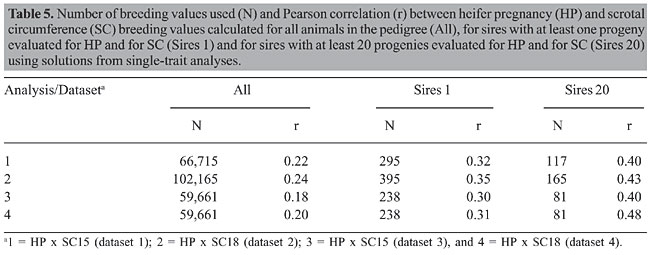
HP and SC data were analyzed simultaneously in four two-trait analyses. Analyses 1, 2, 3, and 4 included records of HP plus records of SC of datasets 1, 2, 3, and 4, respectively. Variance components were estimated using method  procedures with a continuous and categorical animal model (Reverter et al., 1994; Kaiser, 1996). Method  procedures used random 50% sub-samples from the data and compared those sub-samples with all the data. The number of sub-samples used was such that the standard error of the mean heritability was less than 0.0099 for all analyses. The model used for analysis of scrotal circumference (Trait 1) and heifer pregnancy (Trait 2) together can be described as follows (Evans et al., 1999). For SC the model was: and for heifer pregnancy: where y1 = observation of SC, y2 = observation of HP on the liability scale, X1(2) = known coefficient matrix including the incidence matrix relating fixed effects to observations of trait 1(2), Z1(2) = incidence matrix relating random animal additive direct genetic effects to observations of trait 1(2), b1(2) = vector of fixed effects related to trait 1(2), µ1(2) = vector of random additive direct genetic effects for trait 1(2), e1 = vector of random errors associated with trait 1, E (e2|e1) = expectation of errors associated with trait 2 given the observed errors for trait 1, e2* = vector of random errors associated with trait 2 independent of the random errors for trait 1, and b1 = partial regression coefficient for e2 on e1 (Kaiser and Golden, 1995; Kaiser, 1996). Because SC and HP were observed in separate animals, a zero value was assigned to b1. The matrix g = 2 x 2 additive direct genetic (co)variance, A = Wright’s numerator relationship matrix, Ä = Kronecker product, I1(2) = identity matrix of the same order as e1(2), and s2e1 = variance of the SC residual error. The matrix corresponding to the continuous-threshold residual covariance was set to zero because the traits were measured in different animals (Kaiser and Golden, 1995; Kaiser, 1996). Model and fixed effects The model included as fixed effects for HP, the age of the heifer (day of year born) as a linear covariate, and contemporary group. The heifers were distributed among 68 contemporary groups (CG) defined as heifers born in the same herd-year and exposed to breeding in the same herd-year with the same pasture service sire or multi-sire group. Service sires were almost always a multi-sire pasture group which determined the heifer breeding management group. For SC, the model included day of year born as linear and quadratic covariate and fixed effect of CG. CG was defined as bulls born in the same herd-year and evaluated on the same weaning and post-weaning management groups. Only CG with 5 or more animals was kept in the datasets. Animal additive genetic effect and residual error were included as random effects. Pedigree data included all animals with an observation of HP or SC plus all known relationships up to nine generations. The pedigree included 66,715, 102,165, 59,661, and 59,661 animals for datasets 1, 2, 3, and 4, respectively. Genetic evaluation for heifer pregnancy and scrotal circumference For HP, breeding values were predicted based on the underlying scale using a maximum a posteriori probit threshold model - MAP (Gianola and Foulley, 1983; Harville and Mee, 1984; Evans et al., 1999). Solutions were obtained for all animals in the pedigree file using a single-trait animal model and previously reported heritability. EPD were computed by transforming MAP solutions for deviations from 50% probability according to the equation EPDi = [F (MAPi * 0.5) - 0.5] * 100 (Snelling et al., 1995), where: EPDi = expected progeny difference for the ith animal on the probability scale; F = standard accumulated distribution function; MAPi = solution for the ith animal in the underlying scale. Using this transformation, predictions were presented as the probability of a bull having daughters that will become pregnant when exposed at 14 months of age during a given breeding season. For SC, breeding values were predicted using a single-trait linear animal model. Pearson correlation was calculated between HP and SC, for three groups of animals in each of the four datasets: a) for all animals in the pedigree file, b) for sires with at least one progeny simultaneously evaluated for HP and SC and c) for sires with at least 20 progenies evaluated for the two traits simultaneously. The Animal Breeder’s Toolkit software (Golden et al., 1992) was used for variance component estimation and to assemble and solve mixed model equations. Furthermore, 90% confidence intervals of heritability estimates were generated using a procedure described by Mallinckrodt et al. (1997). RESULTS AND DISCUSSION Fixed effects The estimated effect of heifer age on heifer pregnancy was 0.88%/day. This means that for every month increase in heifer age there was a corresponding 26% increase in the probability a heifer would conceive. For SC adjusted to 450 days, the estimated effect of the same covariate was 0.04 (linear effect) and -0.0001 (quadratic). For SC adjusted to 550 days, the estimated effect was 0.02 and -0.0001 for linear and quadratic effects, respectively. SC increased according to the period of the year when the animal was born from January to July and decreased thereafter up to December. As expected, this effect was less important for SC taken at around 18 months of age compared to 15. Heritability and genetic correlation Heritability and genetic correlation estimates from the two-trait analyses are presented in Tables 3 and 4, and the distributions of estimates are presented in Figures 1, 2 and 3. Pearson correlation between HP and SC breeding values are presented in Table 5.
For HP, the mean heritability estimates were similar in the four analyses with values ranging from 0.66 to 0.67 and with standard error from 0.08 to 0.09. Nevertheless, the better 90% confidence interval was found in analysis 2 (0.54 to 0.78). The mean heritability estimates for SC ranged from 0.53 (analysis 2) to 0.64 (analysis 4) and the standard deviation ranged from 0.03 (analysis 2) to 0.07 (analysis 3). The better 90% confidence interval also came from analysis 2 (0.49 to 0.58). When analyzing SC data from animals measured at both ages, 15 and 18 months (analyses 3 and 4), a tendency was seen toward a higher heritability for SC18 (0.64) than for SC15 (0.60). However, analysis 2 which included animals whose records were taken at 18 months (36,091 young bulls) gave lower heritability (0.53) than analysis 1, which included 13,742 records taken at 15 months (0.57). Heritability estimates of 0.52 for this population were reported by Eler et al. (1996; 2001) in analyses using maximum likelihood methods and a two-trait animal model including SC and weaning weight. Based on the standard errors and confidence intervals, h2 estimated from analysis 2 (SC18) seems to be the best one. This estimation can be, however, due to the larger amount of data and better pedigree structure for this specific dataset, which means that heritability estimates derived from analyses 1, 3 and 4 could be overestimated. What is important, however, is that, from these results, it can be concluded that SC and HP are highly heritable traits in this Nellore population whether SC is taken at 15 or 18 months of age. The genetic correlation estimates ranged from 0.11 ± 0.11 (analysis 3) to 0.27 ± 0.10 (analysis 2). As for h2 estimation, the genetic correlation estimate was slightly higher in analysis 4 (SC18) than in analysis 3 (SC15). The estimate was almost twice as high from analysis 2 than from analysis 1. Also in this case, based on the standard errors and confidence intervals, it seems that the genetic correlation estimated from analysis 2 is the best one, but it can be, at least in part, due to the amount of data and pedigree used in this analysis and not only due to age at SC measurement. What is clear from these results is that because SC and HP were observed in separate animals, the genetic correlation can only be well estimated with a large amount of data and well-connected pedigree information, since the estimation comes only from the relationship matrix. In contrast to the h2 estimation, the values obtained from analyses 1, 3 and 4 would be underestimated. Although the value of 0.27 is consistent with the results reported by Martínez-Velázquez et al. (2003) who analyzed data from Bos taurus breeds, the genetic correlation between these two traits can even be higher when analyzing a large dataset with a very good pedigree structure. Results presented in Table 5 show that Pearson correlation between HP and SC breeding values calculated from all the animals in the pedigree file was similar to the estimates from two-trait analyses. However, when only sire breeding values were used for its calculation, the correlation was much higher, reaching up to 0.48 for sires with 20 or more progenies. There was no clear evidence indicating at which age SC data should be collected, but the correlation was always slightly higher in the analyses involving SC collected at around 18 months of age compared to 15 months. One more point that would favor collecting SC data at 18 months of age instead of 15 months is that SC18 was less influenced by the month of birth (or by day of year born). These results, no mater if the correlation is 0.27 or 0.48, still indicate that response to selection for heifer precocity based on SC would be much slower than previously believed for Nellore cattle. On the other hand, as pointed out by Eler et al. (2004), a genetic correlation of such magnitude would be high enough to encourage the incorporation of SC in a two-trait analysis in order to increase accuracy of predicting HP, at least, in young bulls. CONCLUSIONS The high heritability estimates obtained in the present study, for both traits, HP and SC, confirm the use of HP EPD to select for sexual precocity in the Nellore breed. The low estimated genetic correlation between HP and SC indicates that selection based on HP EPD will be more effective than selection based on SC EPD. The results, although not conclusive, indicate that SC measured at around 18 months of age correlates genetically with HP more than does SC measured at around 15 months of age. ACKNOWLEDGMENTS The authors acknowledge financial support from Fundação de Apoio à Pesquisa do Estado de São Paulo (FAPESP), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Agro-Pecuária CFM Ltda. REFERENCES Brinks JS, McInerney MJ and Chenoweth PJ (1978). Relationship of age at puberty in heifers to reproductive traits in young bulls. Proc. West. Sect. Am. Soc. Anim. Sci. 29: 28-30. CFM (2005). Sumário de Touros Nelore. Agro-Pecuária CFM Ltda., Equipe Técnica GMA/USP, São José do Rio Preto e Pirassununga, SP, Brazil. Doyle SP, Golden BL, Green RD, Mathiews GL, et al. (1996). Genetic parameter estimates for heifer pregnancy rate and subsequent rebreeding rate in Angus cattle. J. Anim. Sci. 74: 117 (Abstract). Eler JP, Ferraz JBS and Silva PR (1996). Parâmetros genéticos para peso, avaliação visual e circunferência escrotal na raça Nellore, estimado por modelo animal. Arq. Bras. Med. Vet. Zootec. 48: 203-213. Eler JP, Ferraz JBS and Dias F (2001). Estimação de parâmetros genéticos para características produtivas na raça Nellore. II. Fase pós-desmama. Anales de la XVII Reunion de la Associacion Latino-Americana de Production Animal, Havana, Cuba. (CD-ROM), serie G-3. Eler JP, Silva JA, Ferraz JBS, Dias F, et al. (2002). Genetic evaluation of probability of pregnancy at 14 months for Nellore heifers. J. Anim. Sci. 80: 951-954. Eler JP, Silva JA, Evans JL, Ferraz JBS, et al. (2004). Additive genetic relationships between heifer pregnancy and scrotal circumference in Nellore cattle. J. Anim. Sci. 82: 2519-2527. Evans JL, Golden BL, Bourdon RM and Long KL (1999). Additive genetic relationship between heifer pregnancy and scrotal circumference in Hereford cattle. J. Anim. Sci. 77: 2621-2628. Fajersson P, Barradas HV, Roman-Ponce H and Cook RM (1991). The effects of dietary protein on age and weight at the onset of puberty in Brow Swiss and Zebu heifers in tropics. Theriogenology 35: 845-855. Gallant AR and Fuller WA (1973). Fitting segmented polynomial regression models whose join points have to be estimated. J. Am. Stat. Assoc. 68: 140-147. Gianola D and Foulley JL (1983). Sire evaluation for ordered categorical data with a threshold model. Genet. Sel. Evol. 15: 201-224. Golden BL, Snelling WM and Mallinckrodt CH (1992). Animal breeder’s tool kit user’s guide and reference manual. Tech. Bull., LTB92-2. Colorado State Univ. Agric. Exp. Sta, Fort Collins, CO, USA, 75. Harville DA and Mee RW (1984). A mixed-model procedure for analyzing ordered categorical data. Biometrics 40: 393-408. Kaiser CJ (1996). Incorporating birth weight information into a calving ease threshold model analysis. Ph.D. dissertation, Colorado State University, Fort Collins, CO, USA. Kaiser CJ and Golden BL (1995). Modified equations for analysis of continuous and ordered categorical data with missing observations. Proc. West. Sect. Am. Soc. Anim. Sci. 46: 209-212. Lôbo RB, Bezerra FLA, Oliveira HN, Magnabosco CU, et al. (2005). Avaliação genética de touros e matrizes da raça Nelore. Sumário 2005. ANCP, Ribeirão Preto, SP, Brazil, 28. Mallinckrodt CH, Golden BL and Reverter A (1997). Approximate confidence intervals for heritability from method R estimates. J. Anim. Sci. 75: 2041-2046. Martin LC, Brinks JS, Bourdon RM and Cundiff LV (1992). Genetic effects on beef heifer puberty and subsequent reproduction. J. Anim. Sci. 70: 4006-4017. Martínez-Velázquez G, Gregory KE, Bennett GL and Van Vleck LD (2003). Genetic relationships between scrotal circumference and female reproductive traits. J. Anim. Sci. 81: 395-401. Martins Filho R and Lôbo RB (1991). Estimates of genetic correlations between scrotal circumference and offspring age at first calving in Nellore cattle. Rev. Bras. Genet. 14: 209-212. Mourão GB, Mattos EC, Balieiro JCC, Figueiredo LGG, et al. (2004). Uso de polinômio segmentado para ajuste do peso à desmama da raça Nelore e seu efeito sobre as estimativas de parâmetros genéticos. Anais da Sociedade Brasileira de Zootecnia, 41, 2004. CD-ROM (Melhoramento), Campo Grande, MS, Brazil. Nogueira GP (2004). Puberty in South American Bos indicus (Zebu) cattle. Anim. Reprod. Sci. 82-83: 361-372. PAINT (2005). Sumário consolidado. Equipe técnica PAINT, Lagoa da Serra, Sertãozinho, SP, Brazil, 60. Pereira E, Eler JP and Ferraz JBS (2000). Correlação genética entre perímetro escrotal e algumas características reprodutivas na raça Nelore. Rev. Bras. Zootec. 29: 1676-1683. Pereira E, Eler JP and Ferraz JBS (2002). Análise genética de características reprodutivas na raça Nelore. Pesqui. Agropecu. Bras. 37: 703-708. Quirino CR and Bergmann JAG (1998). Heritability of scrotal circumference adjusted and unadjusted for body weight in Nellore bulls, using univariate and bivariate animal models. Theriogenology 49: 1389-1396. Reverter ABL, Golden R, Bourdon M and Brinks JS (1994). Method R variance components procedure: application of the simple breeding value model. J. Anim. Sci. 72: 2247-2253. Silva JA, Dias JL and Albuquerque LG (2005) Estudo genético da precocidade sexual de novilhas em um rebanho Nelore. Rev. Bras. Zootec. 34: 1568-1572. Smith BA, Brinks and Richardson JS (1989). Relationships of sire scrotal circumference to offspring reproduction and growth. J. Anim. Sci. 67: 2881-2885. Snelling WM, Golden BL and Bourdon RM (1995). Within-herd genetic analyses of stayability of beef females. J. Anim. Sci. 73: 993-1001. |
|