![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
ABSTRACT. The availability of the complete genome of the Gram-negative b-proteobacterium Chromobacterium violaceum has increasingly impacted our understanding of this microorganism. This review focuses on the genomic organization and structural analysis of the deduced proteins of the chemosensory adaptation system of C. violaceum. C. violaceum has multiple homologues of most chemotaxis genes, organized mostly in clusters in the bacterial genome. We found at least 67 genes, distributed in 10 gene clusters, involved in the chemotaxis of C. violaceum. A close examination of the chemoreceptors methyl-accepting chemotaxis proteins (MCPs), and the deduced sequences of the members of the two-component signaling system revealed canonical motifs, described as essential for the function of the deduced proteins. The chemoreceptors found in C. violaceum include the complete repertoire of such genes described in bacteria, designated as tsr, tar, trg, and tap; 41 MCP loci were found in the C. violaceum genome. Also, the C. violaceum genome includes a large repertoire of the proteins of the chemosensory transducer system. Multiple homologues of bacterial chemotaxis genes, including CheA, CheB, CheD, CheR, CheV, CheY, CheZ, and CheW, were found in the C. violaceum genome. Key words: Chromobacterium violaceum, Chemotaxis, Methyl-accepting chemotaxis proteins, Chemosensory transducer protein GENERAL CONSIDERATIONS Bacteria have developed sophisticated signaling systems to allow adaptive responses to the environment. In the chemotaxis network, the receptor and kinase functions are separated in order to allow the cell to execute an integrated response to multiple stimuli (Tsang et al., 1973). The chemosensory system is essential for temporal sensing and for the accumulation of bacteria in environments that are optimal for growth. Chemotaxis begins with the binding of molecules to four kinds of receptors in the plasma membrane. The chemoreceptors are encoded by the tsr, tar, trg and tap genes, also called genes coding for methyl-accepting chemotaxis proteins (MCPs), which are reversibly methylated. These proteins are the sites of initial signal transduction, and they function as homodimers (Falke et al., 1997; Mowbay and Sandgreen, 1998). The MCPs present a cytoplasmic domain that is highly conserved across species (Morgan et al., 1993). MCPs also present conserved glutamate residues, which are methylated during adaptation by a constitutively active methyltransferase named CheR (Stock et al., 1984) and removed by the methylesterase, CheB (Yonekawa et al., 1983). CHROMOBACTERIUM VIOLACEUM METHYL-ACCEPTING CHEMOTAXIS PROTEIN GENES Chromobacterium violaceum is a Gram-negative b-proteobacterium, highly abundant in the water of rivers in Brazil (Caldas, 1990). Recently, the complete genome of the bacteria has been published by a Brazilian National Genome Project Consortium (Ribeiro de Vasconcelos et al., 2003). Here, we briefly review the chemotaxis and flagella genes of C. violaceum, and we describe the organization of the genes in this bacteria’s genome. Forty-one individual open reading frames (ORFs) coding for receptor functions and 26 ORFs coding for proteins that respond to environmental substrates were found in C. violaceum (Table 1). The number of ORFs coding for chemosensing and chemotaxis (n = 67) appears to be high when compared to other bacterial genomes. For example, the genome of Pseudomonas aeruginosa, which has a complex chemosensory system, has 43 ORFs involved in environmental responses, representing 0.8% of the total genome (Stover et al., 2000).
The 41 MCP genes in C. violaceum are found in the four known classes (I, II, III and IV), designated respectively as tsr, tar, trg and tap. Sequences encoding for 13 tsr, 15 tar, 9 trg, and 4 tap were found in C. violaceum (Table 1). These numbers are in agreement with descriptions in the literature. The abundance of the chemotaxis-specific receptors varies among species; tsr and tar are more prevalent than tap and trg (Hazelbauer and Harayama, 1983). The 41 sequences encoding MCPs are considered to be a high number, when compared to many other bacteria. For example, Caulobacter crescentus has 18 MCPs (Nierman et al., 2001) and Halobacterium species NRC-1 has 17 (Ng et al., 2000). The number detected in C. violaceum is similar to that known for Vibrio cholerae (Heidelberg et al., 2000), indicating that C. violaceum has a great ability to respond to a wide variety of compounds. All the MCPs in C. violaceum have at least one transmembrane domain, which enable the proteins to anchor to the membrane and perform a receptor function (data not shown). The MCPs, despite their diversity, have similar domain architectures, with conserved domains (Galperin et al., 2001). The plasmid achromobacter secretion domain, commonly found in the sensor module (Taylor and Zhulin, 1999) of the chemotaxis network, was found in four MCPs of C. violaceum. In addition, the HAMP domain (histidine kinase adenylyl cyclase MCP and phosphatase) that follows the second transmembrane segment, and is putatively related to MCP dimerization (Aravind and Ponting, 1999), was detected in 26 of the 41 MCPs of C. violaceum. Also, the Hpt motif (histidine-containing phosphotransfer), a phosphoacceptor domain, was found in one of the deduced MCPs of C. violaceum (Matsushika and Mizuno, 1998). CHROMOBACTERIUM VIOLACEUM CHEMOSENSORY TRANSDUCER PROTEINS Chromobacterium violaceum appears to have a very complex chemosensory transducer protein system. Twenty-six ORFs involved in the chemotaxis transduction pathway were found in this bacterium. Multiple copies of those genes were found: being four CheA, three CheB, two CheD, three CheR, three CheV, five CheY, one CheZ, and five CheW (Table 1). The number of related genes is identical, and the genes have similar sizes when compared, for example, to the ubiquitous environmental bacterium P. aeruginosa in which 26 ORFs encoding putative chemotaxis transducer proteins were identified (Stover et al., 2000). Those genes in C. violaceum are present mostly in three clusters (Figure 1). 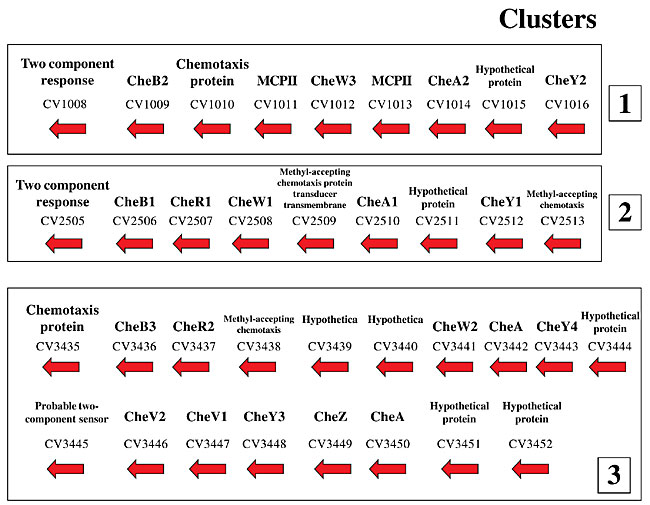
Figure 1. Physical map of the chemotaxis genes in Chromobacterium violaceum. The numbers refer to the ORFs in the C. violaceum genome
(http://www.brgene.lncc.br).
The adaptive response, referred to as a two-component regulatory system (Hock and Silhavy, 1995), often involves at least two components of the signal transduction proteins (Parkinson and Kafoid, 1992), consisting typically of the sensory kinase (CheA) and the response regulator (CheY). Molecular communication between sensors and regulators involves phosphotransfer reactions of the histidine-asparagine phosphorelay (Inouye, 1996). The chemotaxis sensor system networks in C. violaceum have the sensor kinase CheA, and the two-response regulator CheY, as well as additional loci: chew, cheb and chev. The deduced CheA proteins of C. violaceum that could perform the transmitter function present an invariant histidine residue in the P1 domain, which could be autophosphorylated in an ATP-dependent manner, as has been described in other bacteria (Stock et al., 1990; Porter and Armitage, 2002). All the four deduced CheA proteins in C. violaceum also presented the canonical regions of the P4 domain, N, G1, F and G2 (Figure 2). Homology search analysis revealed a high degree of conservation of CheA of C. violaceum and sequences described in the database. For example, identity values ranging from 46 to 61% were found between the C. violaceum CheA and the homologues in E. coli. 
Figure 2. Excerpt of alignment of the deduced CheA proteins of Chromobacterium violaceum and homologues in E. coli. Arrow indicates the phosphorylation site at the P1 domain; N, F, G1 and G2 represent the conserved regions in the P4 domain. The accession number for E. coli CheA is NP_754195.
Changes in chemoreceptor occupancy modulate the kinase activity of CheA in bacteria, which in turn controls the concentration of phosphorylated CheY and CheB. The return of the cell to prestimulation behavior is performed by the methyltransferase CheR, an enzyme that transfers methyl groups from S-adenosylmethionine to glutamate residues on the cytoplasmic domains of the MCPs (Springer and Koshland Jr., 1977; Wu et al., 1996). CheY is a small single-domain protein, a response regulator, which after phosphorylation binds to the FliM component of the flagellar motor switch, inducing clockwise motor rotation (Welch et al., 1993). The deduced CheY of C. violaceum has five amino acid residues (Figure 3) that are conserved among response regulators: D, at positions 12, 13 and 57; T at position 87, and K at position 109, considering the E. coli homolog. The conserved aspartic residue 57 could be the site of phosphorylation, as previously described (Silversmith et al., 2003). Several copies of CheY also occur in other organisms, such as R. sphaeroides (Shah et al., 2000) and Rhizobium mellioti (Greck et al., 1995). 
Figure 3. Alignment of the Chromobacterium violaceum deduced CheY and the homologue in E. coli. Single arrow indicates the phosphorylation site; double arrows indicate the conserved residues of the response regulator.
CheB acts in conjunction with CheR, a methyltransferase, and both are involved in the adaptation process. CheR is responsible for the reversible methylation of the MCPs and CheB in the demethylation process. The CheB homologues present in C. violaceum have a canonical amino-terminal domain, as well as five conserved residues also present in CheY. All of them present the three conserved residues of the catalytic site Ser164, His190 and Asp286, and the GXGXXG nucleotide-binding-fold consensus sequences from other CheB (data not shown; West et al., 1995). The function of CheW is not yet totally clear. This protein is essential for the formation of the ternary complex, CheA-CheW-MCP. CheV is a composite protein that possesses the amino-terminal sequence of CheW and the carboxy-terminal sequence of CheY. CheD has been found in chemotaxis-like operons of a large number of bacteria and archaea, suggesting that it plays an important role in chemotactic sensory transduction in many organisms. However, the molecular function of CheD remains unknown. Kristich and Ordal (2002) demonstrated that CheD catalyzes amide hydrolysis of specific glutaminyl side chains of the B. subtilis chemoreceptor McpA. CheD mutant cells do not respond to most chemoattractants, suggesting that this protein is required for B. subtilis chemoreceptors to effectively transduce signals to CheA kinase. CONCLUDING REMARKS Flagella serve primarily as locomotory organelles in flagellated bacterial species. The chemotactic response is accomplished by signal transmission between the receptor complexes and the flagellar motor complexes. CheY, acting as a messenger protein, transduces the signal from the MCPs to the flagella (Bren and Eisenbach, 2000). Bacteria efficiently respond to changes in the chemical composition of their environment, and this behavior is achieved by integrating the signals received from MCPs, modulating the direction of the flagellar rotation, and hence the swimming direction (Falke et al., 1997). There are around 67 predicted flagellar genes in C. violaceum; 62 were found in seven clusters (Figure 4). The flagellar-motor supramolecular complex at the base of the flagellum includes the proteins FliG, F, M, and N. MotA and MotB form a proton channel anchored to the cell wall (Bren and Eisenbach, 2000). All these proteins were found in the C. violaceum genome. The regulation of expression of flagellar genes seems to have an intricate pattern in many bacteria. The principal regulators are the FlhDC master operon genes, the anti-sigma factor gene FlgM, and the s28 gene FliA (Helmann, 1991). On the basis of sequence comparisons, C. violaceum lacks FlhDC, but has one copy of each the regulators, flgM and fliA, suggesting that the expression of the flagellar genes is under the control of those proteins. 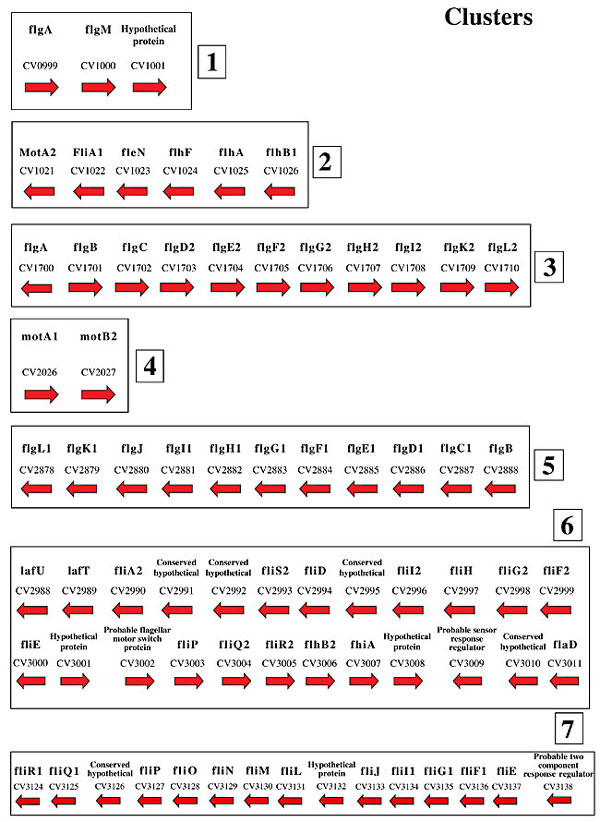
Figure 4. Physical map of the flagellar genes of Chromobacterium violaceum. The numbers refer to the ORFs in the C. violaceum genome
(http://www.brgene.lncc.br).
The finding of 41 MCP genes in C. violaceum (encoding for 13 tsr, 15 tar, 9 trg, and 4 tap), added to the multiple copies of the chemotaxis genes (four CheA, three CheB, two CheD, three CheR, three CheV, five CheY, five CheW, and one CheZ), lead us to suggest that the chemotaxis signal transduction pathway is complex in this free-living microorganism. Investigations into the role of the multiple proteins in the behavior of C. violaceum should be useful to provide insights regarding this bacterium’s adaptations to chemotactic stimuli. ACKNOWLEDGMENTS The study described here was undertaken within the context of the Brazilian National Genome Program (a consortium funded in December 2000 by the Ministério da Ciência e Tecnologia (MCT) through the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). REFERENCES Aravind, L. and Ponting, C.P. (1999). The cytoplasmic helical linker domain of receptor histidine kinase and methyl-accepting proteins is common to many prokaryotic signaling proteins. FEMS Microbiol. Lett. 176: 111-116. Bren, A. and Eisenbach, M. (2000). How signals are heard during bacterial chemotaxis:protein-protein interactions in sensory signal propagation. J. Bacteriol. 182: 6865-6873. Caldas, L.R. (1990). Um pigmento nas águas negras. Ciênc. Hoje 11: 56-67. Falke, J.J., Bass, R.L., Butler, S.L., Chervitz, S.A. and Danielson, M.A. (1997). The two-component signaling pathway of bacterial chemotaxis: a molecular view of signal transduction by receptors, kinases and adaptation enzymes. Annu. Rev. Cell Dev. Biol. 13: 457-512. Galperin, M.Y., Nikolskaya, A.N. and Koonin, E.V. (2001). Novel domains for the prokaryotic two-component signal transduction systems. FEMS Microbiol. Lett. 203: 11-21. Greck, M., Plazer, J., Sourjik, V. and Schimitt, R. (1995). Analysis of chemotaxis operon in Rhizobium mellioti. Mol. Microbiol. 15: 989-1000. Hazelbauer, G.L. and Harayama, S. (1983). Sensory transduction in bacterial chemotaxis. Int. Rev. Cytol. 81: 33-70. Heidelberg, J.F., Eisen, J.A., Nelson, W.C., Clayton R.A., Gwinn, M.L., Dodson, R.J., Haft, D.H., Hickey, E.K., Peterson, J.D., Umayam, L., Gill, S.R., Nelson, K.E., Read, T.D., Tettelin, H., Richardson, D., Ermolaeva, M.D., Vamathevan, J., Bass, S., Qin, H., Dragoi, I., Sellers, P., McDonald, L., Utterback, T., Fleishmann, R.D., Nierman, W.C., White, O., Salzberg, S.L., Smith, H.O., Colwell, R.R., Mekalanos, J.J., Venter, J.C. and Fraser, C.M. (2000). DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae. Nature 406: 477-483. Helmann, J.D. (1991). Alternative sigma factors and the regulation of flagellar gene expression. Mol. Microbiol. 5: 2875-2882. Hock, J.A. and Silhavy, T.J. (1995). Two-Component Signal Transduction. American Society for Microbiology, Washington, DC, USA. Inouye, M. (1996). His-Asp phosphorelay; two components or more? Cell 85: 13-14. Kristich, C.J. and Ordal, G.W. (2002). Bacillus subtilis CheD is a chemoreceptor modification enzyme required for chemotaxis. J. Bacteriol. 277: 25356-25362. Matsushika, A. and Mizuno, T. (1998). The structure and function of the histidine-containing phosphotransfer (Hpt) signaling domain of the Escherichia coli AraB sensor. J. Biochem. 124: 440-445. Morgan, D.G., Baumgartner, J.B. and Hazelbauer, G.L. (1993). Proteins antigenically related to methyl- accepting chemotaxis proteins of Escherichia coli detected in a wide range of bacterial species. J. Bacteriol. 175: 133-140. Mowbay, S.L. and Sandgreen, M.O. (1998). Chemotaxis receptors: a progress report on structure and function. J. Struct. Biol. 124: 257-275. Ng, W.V., Kennedy, S.P., Mahairas, G.G., Berquist, B., Pan, M., Shukla, H.D., Lasky, S.R., Baliga, N.S., Thorsson, V., Sbrogna, J., Swartzell, S., Weir, D., Hall, J., Dahl, T.A., Welti, R., Goo, Y.A., Leithauser, B., Keller, K., Cruz, R., Danson, M.J., Hough, D.W., Maddocks, D.G., Jablonski, P.E., Krebs, M.P., Angevine, C.M., Dale, H., Isenbarger, T.A., Peck, R.F., Pohlschroder, M., Spudich, J.L., Jung, K.W., Alam, M., Freitas, T., Hou, S., Daniels, C.J., Dennis, P.P., Omer, A.D., Ebhardt, H., Lowe, T.M., Liang, P., Riley, M., Hood, L. and DasSarma, S. (2000). Genome sequence of Halobacterium species NRC-1. Proc. Natl. Acad. Sci. USA 97: 12176-12181. Nierman, W.C., Feldblyum, T.V., Laub, M.T., Paulsen, I.T., Nelson, K.E., Eisen, J.A., Heidelberg, J.F., Alley, M.R., Ohta, N., Maddock, J.R., Potocka, I., Nelson, W.C., Newton, A., Stephens, C., Phadke, N.D., Ely, B., DeBoy, R.T., Dodson, R.J., Durkin, A.S., Gwinn, M.L., Haft, D.H., Kolonay, J.F., Smit, J., Craven, M.B., Khouri, H., Shetty, J., Berry, K., Utterback, T., Tran, K., Wolf, A., Vamathevan, J., Ermolaeva, M., White, O., Salzberg, S.L., Venter, J.C., Shapiro, L., Fraser, C.M. and Eisen, J. (2001). Complete genome sequence of Caulobacter crescentus. Proc. Natl. Acad. Sci. USA 98: 4136-4141. Parkinson, J.S. and Kafoid, E.C. (1992). Communication modules in bacterial signaling proteins. Annu. Rev. Genet. 26: 71-112. Porter, S. and Armitage, J.P. (2002). Phosphotransfer in Rhodobacter sphaeroides chemotaxis. J. Mol. Biol. 324: 35-45. Shah, D.H., Porter, S.L., Harris, D.C., Wadhams, G.H. and Armitage, J.P. (2000). Identification of a forth cheY gene in Rhodobacter sphaeroides and interspecies interaction within the bacterial chemotaxis signal transduction pathway. Mol. Microbiol. 35: 101-112. Silversmith, R.E., Guanga, G.P., Betts, L., Chu, C., Zhao, R. and Bourret, R.B. (2003). CheZ-mediated dephosphorylation of Escherichia coli chemotaxis response regulator CheY: Role for the CheY glutamate 89. J. Bacteriol. 185: 1495-1502. Springer, W.R. and Koshland Jr., D.E. (1977). Identification of a protein methyltransferase as the cheR gene product in the bacterial sensing system. Proc. Natl. Acad. Sci. USA 74: 533-537. Stock, J.B., Clarke, S. and Koshland Jr., D.E. (1984). The protein carboxymethyltransferase involved in Escherichia coli and Salmonella typhimurium chemotaxis. Meth. Enzymol. 106: 310-321. Stock, J.B., Stock, A.M. and Mottonen, J.M. (1990). Signal transduction in bacteria. Nature 344: 395-400. Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D., Warrener, P., Hickey, M.J., Brinkman, F.S., Hufnagle, W.O., Kowalik, D.J., Lagrou, M., Garber, R.L., Goltry, L., Tolentino, E., Westbrock-Wadman, S., Yuan, Y., Brody, L.L., Coulter, S.N., Folger, K.R., Kas, A., Larbig, K., Lim, R., Smith, K., Spencer, D., Wong, G.K., Wu, Z., Paulsen, I.T., Reizer, J., Saier, M.H., Hancock, R.E., Lory, S. and Olson, M.V. (2000). The complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 406: 959-963. Taylor, B.L. and Zhulin, I.B. (1999). PAS domains: internal sensors of oxygen, redox potential, and light. Microbiol. Mol. Biol. Rev. 63: 479-506. Tsang, N., MacNab, R. and Koshland Jr., D.E. (1973). Common mechanism for repellents and attractants in bacterial chemotaxis. Science 181: 60-63. Vasconcelos, A.T.R., Almeida, D.F., Hungria, M., Guimarães, C.T., Antônio, R.V., Almeida, F.C., Almeida, L.G.P., Almeida, R., Gomes, J.A., Andrade, E.M., Araripe, J., Araujo, M.F.F., Astolfi Filho, S., Azevedo, V., Baptista, A.J., Bataus, L.A.M., Baptista, J.S., Belo, A., Berg, C.V.D., Bogo, M., Bonatto, S., Bordingnon, J., Brigido, M.M., Brito, C.A., Brocchi, M., Buryti, H.A., Camargo, A.A., Cardoso, D.D.P., Carneiro, N.P., Carraro, D.M., Carvalho, C.M.B., Cascardo, J.C.M., Cavada, B.S., Chueire, L.M.O., Creczynski-Pasa, T.B., Cunha Junior, N.C., Fagundes, N., Falcão, C.L., Fantinatti, F., Farias, I.P., Felipe, M.S.S., Ferrari, L.P., Ferro, J.A., Ferro, M.I.T., Franco, G.R., Freitas, N.S.A., Furlan, L.R., Gazzinelli, R.T., Gomes, E.A., Gonçalves, P.R., Grangeiro, T.B., Grattapaglia, D., Grisard, E.C., Hanna, E.S., Jardim, S.N., Laurino, J., Leoi, L.C.T., Lima, L.F.A., Loureiro, M.F., Lyra, M.C.C.P., Madeira, H.M.F., Manfio, G.P., Maranhão, A.Q., Martins, W.S., Mauro, S.M.Z., Medeiros, S.R. B., Meissner, R.V., Moreira, M.A.M., Nascimento, F.F., Nicolas, M.F., Oliveria, J.G., Oliveira, S.C., Paixão, R.F.C., Parente, J.A., Pedrosa, F.O., Pena, S.D.J., Pereira, J.O., Pereira, M., Pinto, L.S.R.C., Pinto, L.S., Porto, J.I.R., Potrich, D.P., Ramalho Neto, C.E., Reis, A.M.M., Rigo, L.U., Rondinelli, E., Santos, E.B.P., Santos, F.R., Schneider, M.P.C., Seuanez, H.N., Silva, A.M.R., Silva, A.L.C., Silva, D.W., Silva, R., Simões, I.C., Simon, D., Soares, C.M.A., Soares, R.B.A., Souza, E.M., Souza, K.R.L., Souza, R.C., Steffens, M.B.R., Steindel, M., Teixeira, S.R., Urmenyi, T., Wassen, R., Zaha, A. and Simpson, A.J.G. (2003). The complete genome of Chromobacterium violaceum reveals remarkable and exploitable bacterial adaptability. Proc. Natl. Acad. Sci. USA 100: 11660-11665. Welch, M., Oosawa, K, Aizawa, S. and Eisenbach, M. (1993). Phosphorylation-dependent binding of a signal molecule to the flagellar switch of bacteria. Proc. Natl. Acad. Sci. USA 90: 8787-8791. West, A.H., Martinez-Hackert, E. and Stock, A.M. (1995). Crystal structure of the catalytic domain of the chemotaxis receptor methylesterase, CheB. J. Mol. Biol. 250: 276-290. Wu, J.J., Li, J., Long, D.G. and Weiss, R.M. (1996). The receptor binding site for the methyltransferase of bacterial chemotaxis is distinct from the sites of methylation. Biochemistry 35: 4984-4993. Yonekawa, H., Hayashi, H. and Parkinson, J.S. (1983). Requirement of the CheB function for sensory adaptation in Escherichia coli. J. Bacteriol. 156: 1228-1235. |
|