![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
ABSTRACT. Controlled and natural hybridization between cassava and wild relatives does occur. Barriers within the genus appear to be weak due to recent evolution of the group. All Manihot species examined cytogenetically have a chromosome number of 2n = 36. However, they behave meiotically as diploids. The weak interspecific barriers have led to an extremely heterozygous gene pool that may begin a sequence of hybridization followed by speciation. Introgression from cassava into a number of wild species (M. neusana, M. alutacea, M. reptans and M. anomala) has been detected by both morphological marker genes and molecular techniques. Winged fruit, setaceous bracteoles, and wide leaf sinus were dominant genes that came from cassava and appeared in the hybrids. The characteristic protein bands of cassava were recognized in the hybrid seed protein electrophoresis. Key words: Introgression, Cytogenetic behavior, Gene marker, Interspecific hybrids INTRODUCTION Cassava breeders are increasingly interested in the genes of wild relatives, because they offer many opportunities for improving the crop by transferring alien genes. Infertility between these wild relatives and cassava is the limiting factor to make these untapped genetic resources available, so that they can be manipulated in breeding programs. Gene flow from cassava to wild relatives may have played an important role in the evolution of the wild population. The pollen parent in the initial cross may be a cultivated clone or a recent escapee. The opposite pathway is also true. This may involve a wild Manihot plant that functions as the pollen parent in a cross with a cultivated clone, giving rise to a hybrid that backcrosses with wild plants. After several generations have passed the cassava genes are incorporated by the wild species, giving rise to a new type by introgression. Manihot zehntneri Ule is an example of a wild species that may have evolved through this process. The strong morphological resemblance and affinity shown by electrophoresis (Grattapaglia et al., 1986) gives clear evidence for such a relationship. Some factors may favor the hybridization in one or both of the two directions between wild Manihot species and cassava in nature. One of them is that both of these species flower at the same time and for a long period. A second factor is the overlapping of the natural habitat of the wild species and areas of cultivation of cassava, so that vectors can carry pollen between them. Frequently the interspecific hybrid survives, and in many cases it shows vigor and good fitness (Nassar, 1989). I have seen this vigor in several cases of hybridization between cassava and wild Manihot species, such as M. neusana (Figure 1) and M. pohlii. The meiotic division in several cases is normal and pollen fertility is reasonably good (Nassar et al., 1986). This hybrid fertility would enhance the introgression process and the incorporation of alleles from one species to the other. Electrophoresis has clearly revealed gene introgression from cassava into M. zehntneri, M. pilosa and M. glaziovii (Grattapaglia et al., 1986). 
Figure 1. Hybrid of Manihot neusana with cassava.
The progressive introgression between cassava and certain wild species may result in the extinction of the wild species and the creation of a new one, with genes of one parent species being incorporated by the other. The new species, since it is enriched by new genes, will invade new areas and expand its geographic distribution. This happened with M. zehntneri, which was collected from the States of Bahia, Minas Gerais, São Paulo and Mato Grosso do Sul. Manihot reptans and M. cearulescens are also cases of geographic expansion due to introgression (Nassar, 1979, 1982). Manihot zehntneri shows the highest affinity with cassava, based on electrophoresis analysis (Grattapaglia et al., 1986). Morphologically it is difficult to distinguish it from cassava. In controlled crosses it also exhibits high interfertility with cassava (Nassar, 1978b). This case is similar to that reported by Harlan (1961) in Helianthus, and by Dobzhansky (1973) in Iris, Eucalyptus, Penstemon and Tragopogon. All wild Manihot species that have been examined cytogenetically have a chromosome number of 2n = 36 (Nassar, 1978a) (Table 1). Despite this high chromosome number, Manihot species behave meiotically as diploids. Therefore, they are believed to be allopolyploids; this polyploidization seems to have anticipated the emergence of the whole group and it is responsible for their rapid speciation and their weak interspecific barriers, leading to interspecific hybridization. An extremely heterozygous gene pool is thus created, followed by differentiation; this begins a sequence of hybridization followed by speciation. Nassar (1980a) reported frequent hybridization between M. reptans Pax and M. alutacea Rogers et Appan in sympatric natural habitats within which their population boundaries overlap. Morphological marker gene leaf color and bract size were used to identify this interspecific hybridization (Figure 2)

Figure 2. Natural hybrid (right) of Manihot alutacea (left) with Manihot reptans (medium).
Manihot reptans is a typical example of natural hybridization between wild Manihot species. Its range has expanded during the past 100 years (Nassar, 1984), and this is attributed to the continuing gene introgression from other Manihot species. Introgression of M. reptans with germplasm from other species allowed its ecotypes to penetrate and colonize areas where (pure) M. reptans had previously been unable to do so. This phenomenon was also noted in other species, such as M. caerulescens (Nassar, 1980b). From a plant-breeding viewpoint, the value of these hybrids lies in their ability to cross with the cultigen. The marker genes: lobe shape (Figure 3), flower disk color (Figure 4), and fruit shape and color (Figure 5) were discovered in controlled crosses between cassava and wild Manihot species, as well as in natural hybrids between cassava and different species. These genes were used by Nassar (1989) to identify hybridization. Interspecific hybrids of cassava with M. glaziovii, M. neusana, M. anomala, and M. glaziovii were obtained by Nassar (1989, 1995) through controlled crosses, although their frequency was low. The meiotic behavior of several hybrids (cassava with M. neusana and cassava with M. pseudoglaziovii) was studied by Nassar (1991, 1995), who found low hybrid fertility between these species and cassava 
Figure 3. Hybrid of cassava with Manihot anomala.

Figure 5. Fruit shape and color as marker genes in the hybrid.
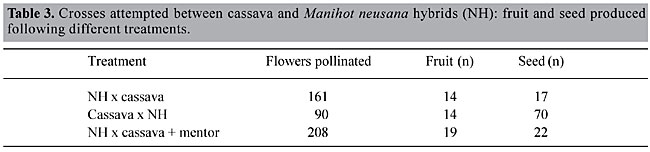
PRODUCTION OF INTERSPECIFIC HYBRIDS BETWEEN CASSAVA AND MANIHOT ANOMALA AND MANIHOT NEUSANA Nassar (1989) reported production of interspecific hybrids of two wild Manihot species, M. neusana Nassar and M. anomala Pax (from the living collection at the Experimental Biology Station, Universidade de Brasília) with cassava through controlled crosses made by vector insects. In October 1982 the wild species were each planted in three rows, alternated with cassava. In June 1983, 200 seeds were collected from each species and replanted in October 1984 for the identification of possible natural hybridization. The following marker genes were used to identify interspecific hybrids: variegated color of fruit dominant to smooth, red color of flower disk dominant to yellow, setaceous bracteole dominant to foliaceous, and noded stem dominant to smooth. Observations of growth habit, height, stem texture, and tuber formation were also recorded. In addition to open pollination of these species, 400 manual crosses with pollen of cassava cultivar Catelo were made. From the 200 M. neusana seeds, only 43 seedlings emerged, and two hybrids were identified. Interspecific hybrids were identified by dominant markers from cassava; noded stem, setaceous bracteoles, ribbed fruit, and tuberculated root. Other characters provided indirect evidence of hybridization. The 200 seeds collected from M. anomala gave rise to 112 seedlings. Among these, three seedlings showed characteristics of interspecific hybridization. Only one of these three survived to maturity. This hybrid plant exhibited dominant phenotypes of cassava, namely ribbed fruit, red color of the flower disk, noded stem, and tuberous roots (Figure 3). Glabrous stem, setaceous-foliaceous bracteoles, red-creamy color of flower disks, variegated-green color of fruit, and ribbed-nonribbed fruit are simple marker genes that can be used to recognize that interspecific barriers between Manihot species can be broken by the use of an abundant diversity of pollinator gametes transmitted by insect vectors. Interspecific crosses were difficult to fertilize manually in this work (Nassar, 1989) and in previous attempts (Nassar et al., 1985a). Barriers between cassava and other Manihot species appear to be weak and recently evolved. It seems they have arisen not as a primary isolating event, but secondarily after geographic isolation. Nassar (1978b) postulated that cassava itself is an interspecific hybrid that appeared by domestication some 200 years ago or less. OVERCOMING BARRIERS BETWEEN CASSAVA AND ITS WILD RELATIVE, MANIHOT POHLII In 1995, Nassar reported results of hybridization of cassava with M. pohlii. Hybrids of M. pohlii and other species with cassava can be obtained, but at a very low frequency due to interspecific crossing barriers (Nassar et al., 1986). However, one technique for overcoming such barriers involves the use of pollen mixes, which combine the pollen intended for syngamy with “mentor” pollen of the maternal species. This technique has been successfully used in a number of plant taxa, including Populus (Knox et al., 1972), Malus (Dayton, 1974), Cosmos (Howlett et al., 1975), and Petunia (Sastri and Shivanna, 1976). The role of the mentor pollen is to facilitate fertilization by foreign pollen. Apparently, the mentor pollen supplies proteinaceous substances that permit the foreign incompatible pollen grains to germinate (Knox et al., 1972). A treatment destroys the generative function of the pollen grain without affecting germination and growth of the pollen tube, and consequently does not affect stimulation. The stimulating effect of destroyed pollen is due to protein recognition substances. They are liberated by pollen grains while germinating, and have enzymatic and antigenic properties. These substances are localized in the internal layer of the pollen surface, and are involved with pollen germination and growth of this tube on the stigma surface. They have been localized by cytochemical means in the cellulose intine (Knox et al., 1972). Many studies using this technique have been successful in overcoming an incompatibility barrier (Brewer and Henstra, 1974). Subsequent work indicated the need to refine the preparation of the mentor pollen by using freeze thaw cycles or methanol treatment (Knox et al., 1972). We report here the successful use of frozen mentor pollen for hybridizing cassava with M. pohlii. In addition to the mentor effect, other techniques seem to have potential for overcoming interspecific barriers. One of these is the use of a bridge species. We were inspired by the capacity of a species, M. neusana Nassar, to cross easily with all Manihot species growing in the vicinity, and have used it in a bridge technique. Probably the only published case of this is Dionne (1963) who used Solanum acaule Bitter as a bridge between Solanum tuberosum L. and S. bulbocostanum Dunal. We report here the use of M. neusana Nassar as a bridge between M. esculenta and M. pohlii. A natural hybrid between M. pohlii and M. neusana Nassar was used as a bridge species. The hybrid was identified by fruit marker genes, which produce variegation of fruit color in M. neusana, and a straight white line in M. pohlii fruit (Figure 5). Crosses of this hybrid (named NHP) and cassava (clone EB05) were carried out beginning in January 1994. Flowers were taped shut for two days until they had been pollinated manually. Pollination of both the hybrid NHP and cassava was done with pollen mixes of cassava and M. pohlii. Manihot pohlii pollen, used as a mentor pollen, was successively frozen for 5 min at -4oC, and thawed for 30 min for a period of 105 min. The purpose of this treatment was to kill the mentor pollen and thereby increase the chance of obtaining interspecific hybrid seed. To determine if there was any auto-incompatibility that would interfere in the controlled crosses between M. pohlii and cassava, a controlled auto-pollination was undertaken. A cross between cassava and M. pohlii was carried out using mentor pollen in one trial and untreated pollen in a second trial. Seeds were collected from both crosses and planted in the next growing season. The pollination of M. pohlii by untreated cassava pollen did not produce any fruit set (Table 2), while crosses with mixed pollen of cassava and mentor resulted in the production of 21 seeds, 4.9% of the possible maximum (assuming that every fruit has three ovules). Using cassava as a maternal parent, pollination by untreated pollen of M. pohlii did not result in any seed. This clearly demonstrates the effect of mentor pollen in the crosses of cassava with M. pohlii. It likewise means that the freeze-thawing treatment administered to M. pohlii pollen, although killing it, did not affect its stimulatory function, so that all of the seed produced by the mentor effect had embryos and endosperm.
Apparently only the stigmatic barrier functions in preventing crossing between these species. Postfertilization mechanisms fail to prevent crossing. Only one plant germinated and could be raised from these seeds to severe dormancy. This plant bears fruit carrying the marker genes of both M. pohlii and cassava; a straight white line from M. pohlii, and winged fruit from cassava (Figure 5). The mentor effect has also been successfully used in Populus and Cosmos. In these genera, interspecific incompatibilities have been overcome by using compatible but dead pollen (Knox et al., 1972). These studies have suggested that this phenomenon is due to proteinaceous recognition factors released from the wall of the killed compatible pollen, masking the rejection reaction of the recipient stigma. This is the first report of obtaining hybrid seed of M. pohlii and cassava, and its further reproduction. In spite of the useful characters of M. pohlii, no successful breeding program has been carried out yet, due to a lack of hybrids between this species and cassava. This is the first successful cross of these two species. The use of M. neusana as a bridge species through the hybrid M. neusana-M. pohlii has improved seed set. When used as a maternal parent pollinated by cassava it gave seed in 3.5% of the cases, while the reciprocal crosses had greatly improved seed set, yielding 25.9% (Table 3). The combined treatment of both species (bridge and mentor) produced 3.5% seed production. The success of M. neusana as a bridge species between M. pohlii and cassava may occur because the two genomes of M. neusana carry different genetic mechanisms of cross incompatibility. This hypothesis is confirmed by our observations on crossing behavior of this species in the living collection of wild Manihot species. Manihot neusana has hybridized naturally with several Manihot species grown in the vicinity, e.g., M. pseudoglaziovii Pax et Hoffm., M. caerulescens Pohl, M. salicifolia Pohl, M. pohlii and M. esculenta itself (Nassar, 1989). Its hybrids with the above mentioned species were easily identified by the dominant marker gene of variegated fruit, which came from M. neusana (Figure 6). Manihot neusana is a recently described and identified species (Nassar, 1985b).

Figure 6. Marker genes of winged fruit (left) and variegated fruit (right).
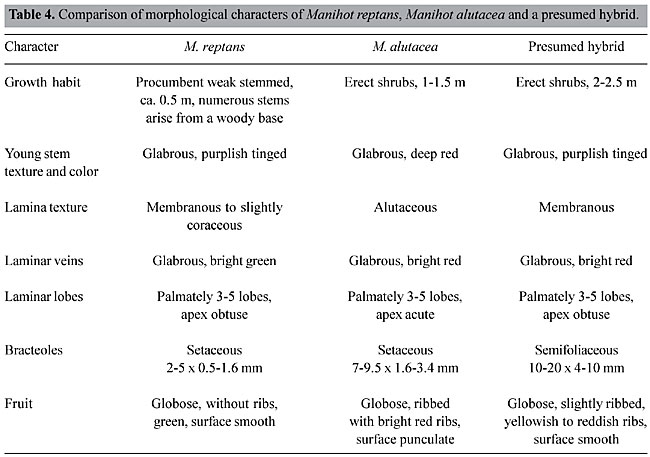
NATURAL HYBRIDIZATION BETWEEN WILD MANIHOT SPECIES Hybridization between wild Manihot species does occur in nature, and it seems to be responsible for rapid speciation in this genus. Natural hybrids between M. reptans and M. alutacea were observed in 1977 (Nassar, 1978c) in two localities in the State of Goiás, where M. reptans grows near M. alutacea, and where these species have a sympatric distribution. Since interspecific hybrids have long been of special interest to breeders because of their heterogeneity, this case has been investigated in further detail. Seeds of M. reptans, M. alutacea and their natural hybrid were collected from two localities in the State of Goiás 10 and 12 km east of the city of Goiás Velho and were planted in a living collection of Manihot at the Universidade de Brasília. Observations of the following characters were recorded: growth habit, young stem, lamina texture and color, laminar veins, laminar lobes, bracteoles and fruit. Thirty bracteoles were measured on each of 10 plants of each species. The morphology of M. reptans, M. alutacea and their hybrid are compared in Table 4. Taxonomically, M. reptans and M. alutacea have been separated on the basis of growth habit, leaf shape and color, plant height, laminar texture and fruit shape and color. The presumed hybrid is similar to one or the other parent for all characters except bracteole size; the bracteoles are much larger than in either parent. Morphological measurements of the presumed hybrid provide indirect evidence of interspecific hybridization. From the scatter diagram (Figure 7), a distinction between bracteole length, range of the two species and their hybrid is seen. Many workers expect the hybrid to occur in areas where distribution of the two parent species adjoins each other and where hybridization between the two is likely to be occurring continuously (Baker, 1951; Anderson, 1953; Heiser, 1961; Harlan and de Wet, 1963; Levin 1983).

Figure 7. Scatter diagram of hybridization of Manihot reptans x Manihot alutacea.
The presumed hybrids that were collected from a site 10 km east of Goiás Velho occurred in the absence of M. reptans stand being swamped by a large amount of pollen from the more abundant M. alutacea. Another possible hypothesis is that both species were present at this site at some early time and hybridized at that time; subsequently the environment changed and M. reptans disappeared, since it was poorly adapted. Our observations indicate that M. reptans and M. alutacea will readily hybridize when grown sympatrically. Sympatric populations contain a large number of hybrids, suggesting absence of barriers that may exclude the hybrids from the gene pool of the population. Nassar (1984) discussed an expansion of the M. reptans range during the last hundred years and attributed this to continuing introgression of genes of Manihot species. The hybrid of M. reptans with M. alutacea was identified by its bright red leaves, a characteristic of M. alutacea that served as a marker gene in this case. A population of M. reptans, with characteristic fruit, and two characteristics of cassava (M. esculenta) was found about 15 km north of Corumbá de Goiás. Apparently, these two characteristics were acquired by M. reptans through introgressive hybridization with cassava. It seems that M. reptans became introgressed with germplasm from other species and was able to extend its ecotypic range and colonize new areas where the pure M. reptans had been previously unable to penetrate. The hybrid type from Corumbá was collected from a disturbed habitat, which may account for its weedy nature. It may be considered part of a weed-crop complex, following the model described by Harlan and de Wet (1963) and others. Hawkes (1977) found these hybrids of great value in plant breeding programs because of their crossability with the cultivated plant. CYTOGENETIC BEHAVIOR OF HYBRIDS OF CASSAVA WITH MANIHOT NEUSANA The wild Manihot species, M. neusana Nassar, was hybridized with the cassava clone Catelo through controlled hybridization, with the help of pollinating insects (Nassar, 1989). An interspecific hybrid that combined marker genes of both parents was obtained. The marker genes were ribbed fruit, acquired from cassava, and variegated fruit color from M. neusana. This hybrid (NH) was backcrossed with cassava and used as a pollinator in the first trial, and as a fruit carrier in the second trial. Seeds were obtained from both crosses, but only one plant could be raised from each; HO1 was the result of the interspecific hybrid (NH) as a maternal plant (seed carrier), and HO4 resulted from crosses in which the interspecific hybrid (NH) was used as a pollinator. The three hybrid plants (NH, HO1 and HO4) were studied cytogenetically for both meiotic and mitotic behavior. For the study of meiosis, inflorescences were fixed in a mixture of three parts absolute alcohol and one part glacial acetic acid and kept in a refrigerator for 24 h. The anthers were smeared with acetic carmine. Chromosome configurations in metaphase, chromosome distribution in anaphase I and tetrad formation were also studied. Pollen viability was determined by using acetocarmine and iodine stain (Nassar, 1978a). For the mitotic study, root tips were left in 0.2% colchicine for 2 h and then fixed in acetic alcohol for 24 h. Before smearing and staining with acetocarmine, they were treated with 1 N HCl for10 min. One hundred pollen mother cells (PMCs) were studied in metaphase I of the interspecific hybrid, M. neusana with cassava; 30 PMCs in metaphase II and 1000 tetrads of the same material were also investigated. The study of metaphase I showed various chromosome configurations (Table 5). The average bivalent frequency in all cells of metaphase I was 16.13 per cell. The high frequency of univalents was attributed to a lack of synapses between chromosomes or failure of the two species to remain associated. Virtually the only other report on this subject is that of Magoon et al. (1970), in which chromosome pairing in the interspecific hybrid M. glaziovii (rubber tree) and cassava was studied and the normal synapses led the authors to conclude that there is a strong relationship between this species and cassava. Nassar et al. (1986) suggested that the material used by Magoon et al. was not pure M. glaziovii, but rather a natural interspecific hybrid between this species and cassava. If this were true, the supposed interspecific hybrid would be backcrossed progeny. The study of anaphase I showed that of 40 PMCs, 38 cells exhibited laggards, which were attributed to the occurrence of univalents, resulting from a lack of homologous chromosomes. 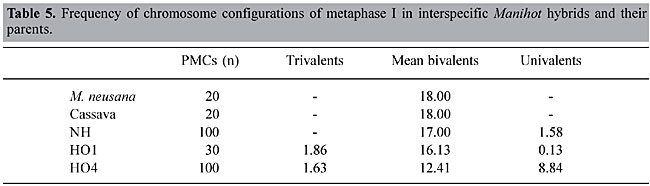
PMC: pollen mother cell
NH: hybrid of cassava with M. neusana HO1: hybrid of NH as a maternal plant (seed carrier) and cassava as a pollinator HO4: NH was used as a pollinator and cassava as a maternal plant Anaphase II showed meiotic restitution. Of 33 PMCs studied in this phase, 5 cells exhibited a second meiotic restitution, forming 36 chromosomes on each pole. Apparently this phenomenon is a consequence of meiotic disturbance in the hybrid. An example of this disturbance was the breakdown of anaphase I. This was probably due to disharmony between the two different genomes. Nassar (1991) documented this phenomenon in the interspecific hybrids of cassava with M. pseudoglaziovii. The presence of such restitution was confirmed in the following tetrad stage, in which the formation of both dyads and tetrads was observed. In various crops, interspecific hybridization has led to the disturbance of meiotic division, with consequent meiotic restitution, e.g., in Trifolium pratense by Parrot and Smith (1984) and Medicago spp. by Vorsa and Bingham (1979). In manioc species, Hahn et al. (1990) reported 2n pollen formation in wild species in addition to certain clones of cassava. The detection of this phenomenon enabled these researchers to isolate triploid and tetraploid types from progeny that came from crosses of cassava with certain wild Manihot species, namely, M. glaziovii and M. epruinosa. These types proved to be much more productive than commercial clones used in Nigeria. Nassar (1991) manipulated the meiotic restitution occurring in the interspecific hybrids of M. pseudoglaziovii with cassava to produce triploid types that showed very good productivity under semiarid conditions. The discovery of the frequent occurrence of this phenomenon in interspecific hybrids of cassava offers an effective tool for the production of polyploid types by sexual means instead of the traditional method of colchicine applied to vegetative parts, which normally induces unstable, chimeral types (Abraham et al., 1964). An additional advantage is that production of triploid types may lead to the production of trisomics among their progeny. If genes that control productivity in cassava are polygenes with an addictive model action, as is the case for many crops certain trisomics of this crop may be more productive than their diploid ancestors. In general, the production of polyploidy via sexual means is advantageous from both genetic and evolutionary standpoints because it offers a vigorous heterotic effect and it releases useful genetic variability adaptations. ACKNOWLEDGMENTS Research supported by the National Council for Scientific Development (CNPq), Brasília. The living collection was established at the Universidade de Brasília with the help of the International Development Research Center (IDRC), Ottawa, to whom this author is grateful. REFERENCES Abraham, A., Panicher, P.K. and Mathew, P.M. (1964). Breeding of tuber crops in India. J. Genet. Plant Breed. 17: 212-217. Anderson, E. (1953). Introgressive hybridization. Biol. Rev. 28: 280-307. Baker, H.G. (1951). Hybridization and natural gene flow between higher plants. Biol. Rev. 5: 302-336. Brewer, J.L. and Henstra, S. (1974). Pollen of Pyrethrum. Fine structure recognition. Euphytica 23: 567-663. Dayton, D.F. (1974). Overcoming self incompatibility in apple with killed compatible pollen. J. Am. Soc. Hort. Sci. 99: 190-192. Dionne, L.A. (1963). Studies of the use of Solanum acaule as a bridge between Solanum tuberosum and species in the series Bulbocastana, Cardiophylla and Pinnatisecta. Euphytica 12: 263-369. Dobzansky, T. (1973). Genética do Processo Evolutivo. Tradução de Celso Adbbade Mourão. Polígono, São Paulo, SP, Brazil. Grattapaglia, D.E., Nassar, N.M.A. and Dianese, J. (1986). Biossistemática de espécies brasileiras do gênero Manihot baseada em padrões de proteína da semente. Cienc. Cult. 19: 294-300. Hahn, S.K., Bai, K.V. and Asiedu, R. (1990). Tetraploids, triploids and 2n pollen from diploid interspecific crosses with cassava. Theor. Appl. Genet. 79: 433-439. Harlan, J. (1961). Geographic origin of plants useful to agriculture. In: Germplasm Resources. Am. Assoc. Adv. Sci. 66: 3-19. Harlan, J.R. and de Wet, J.M. (1963). The compilospecies concept. Evolution 17: 497-501. Hawkes, J.G. (1977). The importance of wild germplasm in plant breeding. Euphytica 26: 615-621. Heiser, C.B. (1961). Natural hybridization and introgression with particular reference to Helianthus. Rec. Adv. Bot. 1: 874-877. Howlett, B.J., Knox, R.B., Paxton, J.H. and Heslop-Harison, J. (1975). Pollen-wall proteins: Physiochemical characterization and role in self-incompatibility in Cosmos bipinnatus. Proc. R. Soc. Lond. 188: 167-182. Knox, R.B., Willing, R.R. and Pryor, L.D. (1972). Interspecific hybridization in Populus using recognition pollen. Silvae Genet. 21: 3-4. Levin, D.A. (1983). Natural hybridization between Phlox maculata and P. glaberrima and its evolutionary significance. Am. J. Bot. 50: 714-719. Magoon, M.L., Krishnan R. and Vijaya Bai, K. (1970). Cytogenetics of the F1 hybrid between cassava and Ceará rubber and its back crosses. Genetica 41: 425-426. Nassar Nagib, M.A. (1978a). Conservation of the genetic resources of cassava: Determination of wild species localities with emphasis on probable origin. Econ. Bot. 32: 311-320. Nassar Nagib, M.A. (1978b). Wild Manihot species of Central Brazil for cassava breeding. Can. J. Plant Sci. 58: 257-261. Nassar Nagib, M.A. (1978c). Some further species of Manihot with potential value to cassava breeding. Can. J. Plant Sci. 58: 915-916. Nassar Nagib, M.A. (1979). Three Brazilian Manihot species with tolerance to stress conditions. Can. J. Plant Sci. 59: 533-555. Nassar Nagib, M.A. (1980a). Attempts to hybridize wild Manihot species with cassava. Econ. Bot. 34: 13-15. Nassar Nagib, M.A. (1980b). The need for germplasm conservation in wild cassava. Indian J. Genet. Plant Breed. 39: 465-470. Nassar Nagib, M.A. (1982). Collecting wild cassava in Brazil. Indian J. Genet. Plant Breed. 42: 405-411. Nassar Nagib, M.A. (1984). Natural hybrids between Manihot reptans Pax and Manihot alutacea Rogers & Appan. Can. J. Plant Sci. 64: 423-425. Nassar Nagib, M.A. (1985a). Genetic variation of wild Manihot species native to Brazil and its potential for cassava improvement. Field Crops Res. 13: 177-184. Nassar, N.M.A. (1985b). Manihot neusana Nassar: A new species native to Paraná, Brazil. Can. J. Plant Sci. 65: 1097-1100. Nassar, N.M.A. (1989). Broadening the genetic base of cassava, Manihot esculenta Crantz by interspecific hybridization. Can. J. Plant Sci. 69: 1071-1073. Nassar Nagib, M.A. (1991). Production of triploid cassava, Manihot esculenta Crantz, by hybrid diploid gametes. Field Crop Res. 13: 173-182. Nassar, N.M.A. (1995). Development of cassava interspecific hybrids for savanna (cerrado) conditions. J. Root Crops 22: 9-17. Nassar, N.M.A., Silva, J.R. and Vieira, C. (1986). Hibridação interespecífica entre mandioca e espécies silvestres de Manihot. Cienc. Cult. 38: 1051-1065. Parrot, W.A. and Smith, R.R. (1984). Production of 2n pollen in red clover. Crop Sci. 24: 467-472. Sastri, D.C. and Shivanna, K.R. (1976). Attempts to overcome interspecific incompatibility in Sesanum by using recognition pollen. Ann. Bot. 40: 891-893. Vorsa, N. and Bingham, E.T. (1979). Cytology of pollen formation in diploid alfalfa, Medicago sativa. Can. J. Genet. Cytol. 21: 526-530. |
|
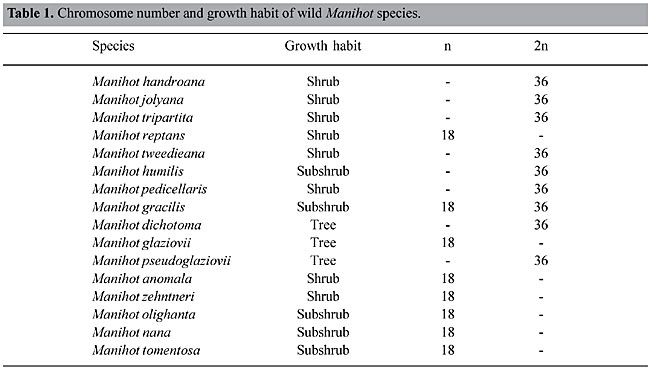
 Figure 4. Flower disk as a marker gene.
Figure 4. Flower disk as a marker gene.