![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
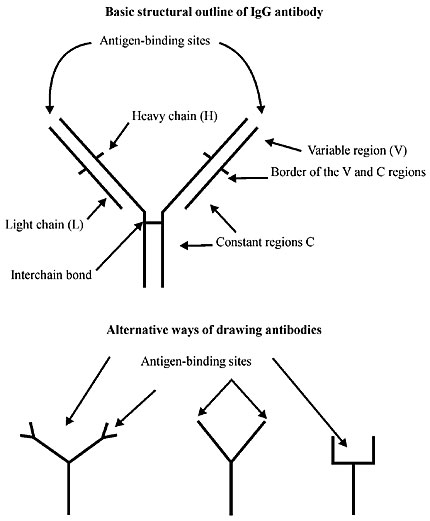
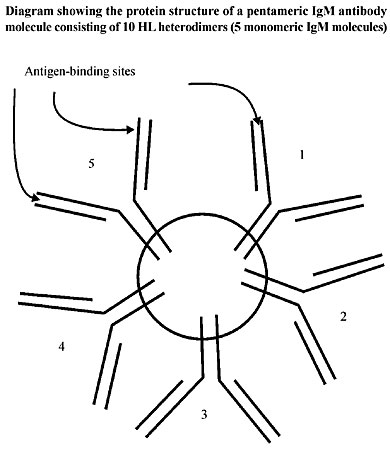
ABSTRACT. The evolvability of vertebrate systems involves various mechanisms that eventually generate cooperative and nonlethal functional variation on which Darwinian selection can operate. It is a truism that to get vertebrate animals to develop a coherent machine they first had to inherit the right multicellular ontogeny. The ontogeny of a metazoan involves cell lineages that progressively deny their own capacity for increase and for totipotency in benefit of the collective interest of the individual. To achieve such cell altruism Darwinian dynamics rescinded its original unicellular mandate to reproduce. The distinction between heritability at the level of the cell lineage and at the level of the individual is crucial. However, its implications have seldom been explored in depth. While all out reproduction is the Darwinian measure of success among unicellular organisms, a high replication rate of cell lineages within the organism may be deleterious to the individual as a functional unit. If a harmoniously functioning unit is to evolve, mechanisms must have evolved whereby variants that increase their own replication rate by failing to accept their own somatic duties are controlled. For questions involving organelle origins, see Godelle and Reboud, 1995 and Hoekstra, 1990. In other words, modifiers of conflict that control cell lineages with conflicting genes and new mutant replication rates that deviate from their somatic duties had to evolve. Our thesis is that selection at the level of the (multicellular) individual must have opposed selection at the level of the cell lineage. The metazoan embryo is not immune to this conflict especially with the evolution of set-aside cells and other modes of self-policing modifiers (Blackstone and Ellison, 1998; Ransick et al., 1996). In fact, the conflict between the two selection processes permitted a Lamarckian soma-to-germline feedback loop. This new element in metazoan ontogeny became the evolvability of the vertebrate adaptive immune system and life as we know it now. We offer the hypothesis that metazoan evolution solved this ancient conflict by evolving an immunogenetic mechanism that responds with rapid Lamarckian efficiency by retaining the ancient reverse transcriptase enzyme (RNA® DNA copying discovered by Temin in 1959 (see Temin, 1989) and found in 1970 in RNA tumor viruses by Temin and Baltimore), which can produce cDNA from the genome of an RNA virus that infects the cells. It seems that molecular Lamarckism can survive (Lewin, 1993). Key words: Evolvability process, Lamarckian selection, Darwinian selection, Cell biology, Molecular embryologyn INTRODUCTION The capacity of a multicellular process to evolve, its evolvability, is principally applied to generating cooperative and nonlethal functional variation, on which selection can operate. However, the evolvability of unicellular processes operates through quite different modes of action whenever conflicting genomes, resulting from mutation, prevail in uniform environments. Prokaryotes have evolved extremely varied metabolic capacities to compensate their quite limited morphological diversification; in fact, prokaryotes have never evolved true multicellularity. Prokaryotes emphasized adaptive success in large populations as a means of statistical survival, while the metazoan evolution record suggests that these organisms diversified and radiated in small populations. Universal characteristics of animal cells, such as their lack of cell walls and their morphological plasticity in epithelial sheets, tubes and cell masses, indicate that their great diversity of forms may have been a prerequisite for the evolution of metazoa. Notwithstanding the conservation of cellular processes and protein function in eukaryotes, there have been extensive changes in DNA and protein sequences, even among morphologically similar species (Gerhart and Kirschner, 1997). We can examine some unusual biochemical properties of cellular and developmental systems generally shared by metazoa, which upon analysis of the properties of these systems has revealed several common principles that may have contributed to the evolvability of organisms from a common stock over the past 600 million years. These devices include flexibility, complex exploratory behavior, developmental robustness, cooperation (cellular and otherwise), and compartmentalization, among others. Cellular and developmental appliances possessing these properties seem to have been selected at different levels (probably at different times) in large part for the physiological and developmental adaptability they provide for the individual organism. In “The Future of Selection” (Hoenigsberg, 2002) we analyzed the various levels of the biological hierarchy: genes, chromosomes, cells, species, and communities, as different survival entities that require different heritable variations in fitness in order to function as units of selection in the evolutionary process. THE MODERN SYNTHESIS AND WEISMANN´S DOCTRINE The classic, though erroneous view, has the zygote producing somatic cells via mitosis, and germ cells via meiosis. Thus, genetic variation arising during the course of ontogeny cannot be inherited. Heritable variation only occurs in the zygote or during the reduction division of gametogenesis. This is Weismann´s doctrine: the germline and the soma were separated to make it possible to understand genetics, and hence Darwinian Evolution, without understanding development. To understand individuality all one had to do was to consider the adult individual as a discrete unit, with heritability limited to a very small group of genetically, more or less homogeneous cells. Multicellular organisms are composites. Individuals are composed of cells capable of division and of variation. Inside eukaryotic cells there are organelles capable of reproduction and variation. In some cases these individuals are units of selection able to condition fitness and differential survival where selfish genes and cooperating gene systems exist as gene sequences in nuclei. Thus, multicellular systems have developed mechanisms to cope with incoming viruses and bacteria that unicellular organisms can avoid. The devastating effect of parasites that enter the multicellular individual can jeopardize reproduction at the onset because only a few cells of the clonemates retain the capacity of giving rise to a new multicellular individual. On the other hand, a unicellular alga dividing by mitosis produces a clone of daughter cells, each of which is capable of giving rise to a new multicellular individual. An encounter with parasites does not have a lethal effect in the latter; other cells of the clone can take over. The irreversible differentiation of cells to purely somatic functions denies a lineage the capacity to produce a new organism and to defend itself from infectious diseases. Thus, it is probable that infectious diseases have been the main, if not the only selective force involved in the evolution of multicellular immune systems. Metazoan evolution solved its fragility by evolving an immunogenetic mechanism that responds with rapid Lamarckian efficiency by retaining the ancient reverse transcriptase enzyme (RNA® DNA copying) discovered by H. Temin (1959, see Temin, 1989), called reverse transcriptase, and found in 1970 in RNA tumor viruses by Temin and Baltimore, of an innocuous retrovirus that can produce DNA copies of the genome of another RNA virus that infects the cells. DOWNLOADING RNA GENETIC BLUEPRINTS TO DNA In 1997 a publication in Nature by the group of Rolf Zinkernagel (Bachmann et al., 1997) showed that certain animal cells that have the reverse transcriptase enzyme of an innocuous retrovirus could produce DNA copies of the genome of another RNA virus that infected the cells. This extraordinary discovery implies that reverse transcriptase enzymes encoded by retroviruses can make DNA copies of other RNA molecules present in the host cell. Thus, the free retrotranscripts (cDNA) of cellular genes could be produced from mRNA templates. In essence this is the soma-to-germline theory (SGT) of evolution for the immune system (proposed by Ted Steele in 1981). The evolutionary process, in terms of the Darwinian survival of the fittest immune system, maintains that infectious diseases have been the principal selective force in the evolution of the vertebrate immune system. If this is true, then the immunoglobulin (Ig) genes, which are responsible for encoding antibodies, are transmitted to progeny via the germ cells and their preservation is dictated by the same Darwinian rules of fitness that apply to other genes. The problem with this general mechanism is that some animals do not possess an adaptive immune system. For example, hagfish and lampreys do not have an adaptive immune system; they do not have Ig variable (IgV) or T cell receptor (TCR) genes (Steele et al., 1998). On the other hand, cartilaginous fish (sharks and rays), that arose at least 400 to 500 million years ago, do have genes corresponding to IgV or TCR genes (Litman, 1996). The main elements of the adaptive immune system of higher warm-blooded vertebrates can even be found in cold-blooded vertebrates (fish). They all have the capacity to: 1. make a very large repertoire of antibodies and T cells; 2. to build an enhanced memory response when a new infection penetrates, and 3. to maintain self-tolerance. It is apparent that in the transition from cold-blooded cartilaginous fish to the warm-blooded land animals the immune system has become more complex. The point we want to make is that for the transition from unicellularity to multicellularity there is an important transition to make. While unicellular organisms have recurred to a reproductive strategy to survive sudden changes in the environment, multicellular organisms had to wait to develop a complex immune system, with genetic variation arising during the course of ontogeny, in order to survive microbial infection. The zygote, by producing somatic cells via mitosis, accumulates a very large probability of getting, among other things, infected. Heredity is not simply a matter of macromolecules, information and code. We must recognize that knowledge of the molecular mechanics of heredity is not equivalent to knowledge of the units which prove to be heritable. Recognition that developmental processes control heritability focuses the issue of inheritance on questions concerning patterns of developmental determinism. If there are only a few cell divisions intercalated between fertilization and the final determination of the germline in each generation, then the opportunity for heritable variation to arise in the course of ontogeny is small and Weismann’s view of the individual as a genetically homogeneous unit is largely vindicated (Buss, 1987). If, however, ontogeny allows embryonic cells to vary and yet still gain access to the gametes, then genetic variation arising during the ontogeny of an individual must be acknowledged as a potentially important source of transmissible variation. In fact, this is the case for organisms with somatic embryogenesis, where a distinct germline is lacking. In these taxa, one cell lineage is capable of somatic function as a stem cell lineage, but it is also competent to give rise to gametes throughout ontogeny (as in plants). This is the case for interstitial cells of Hydra that remain multipotent and mitotically active throughout the life span of the animal (Grayson, 1971; Prasad and Mookerjee, 1986). When local conditions deteriorate, Hydra may be induced to cease asexual reproduction, and instead the interstitial cells differentiate into gametes. However, between each sexual generation a great many number of asexual iterations may occur (Ewer, 1948; Campbell, 1983; Rastogy et al., 1992). In contrast, organisms with preformistic development, such as Drosophila, mice, birds and humans (laboratory animals for the Modern Synthesis) have early embryonic determination of the germline and therefore are animals in which the individual closely approximates a genetically homogeneous unit of selection. The ontogeny of a metazoan generally involves a series of events in which cell lineages progressively deny their own capacity for totipotency to the collective interest of the individual. The question is, how could a taxon evolve such that some cells abandon their own capacity for replication? In fact, that is what happened to some metazoans with preformistic development. However, our point is that it was not the case during their origin. THE SOMA-TO-GERMLINE FEEDBACK Rothenfluh and Steele (1993), Rothenfluh et al. (1995) have provided most of the detailed molecular data for V genes, where DNA sequence structure and germline genes and pseudogenes appear as evidence of what could have happened in the vertebrate immune system. The nonrandom distribution of recombination errors fits expectation of V genes in soma-to-germline feedback, and moreover, exogeneous DNA can be taken up and integrated into genomic DNA and mouse sperm (Spadafora, 1998; Giordano et al., 2000). Moreover, Pollard (1984) discussed Weismann’s doctrine as a mistaken paradigm, but he failed to clarify Weismann’s great contribution, which was the recognition that heritability is controlled by development. Developmental patterns that violate Weismann’s doctrine were well known when Mendelian genetics was recognized as an important element in Neo-Darwinian theory. Steele (1979) was the first to offer a theory of somatic selection and adaptive evolution to explain the importance of the inheritance of acquired characters. Moreover, Steele’s ‘Lamarck and Immunity; a Conflict Resolved’ (1981) demonstrated the intensity of the controversy. We are conscious of the emotive atmosphere that embraces Lamarckism even in todays molecular intellectual milieu. In what follows we intend to apply Lamarckism to resolve the impasse of the evolutionary transition between unicellular and multicellular organisms. After all, the history of life is a history of transitions between different units of selection. The transition we focus on here is the one between the early multifunctional cell and the individual. Other transitions must have preceded it. We maintain that at its origin metazoan ontogeny developed an intensive Darwinian selection in favor of a mechanism that responded to microbial infection. We propose that at that time the mode of development was closer to the model of unicellular algae dividing by mitosis producing a clone of daughter cells in which all descendants were capable of experimenting with transient incipient multicellularity (Xiao et al., 1998). In such early metazoan somatic embryogenesis, although participating with a germline, there was no distinct germline when local conditions deteriorated (as in Hydra), interstitial cells would simply differentiate into gametes. Today we find successful multicellular representatives of the Kingdom Protista (with the exception of the Volvacales) with somatic embryogenic development patterns such as the one we are proposing here. DARWINIAN SELECTION AND THE IMMUNE SYSTEM Organisms are exposed to an inexhaustible and implacable attack of potentially invasive pathogenic or innocuous viruses and bacteria. Innocuousness does not imply that the multicellular animal or plant will not have to biochemically respond to the pervasion. A foreign body brings in another life system, which does not necessarily signify coherence. How does the immune system fight such a multitude of relentless wars, some of which are potentially lethal? Moreover, in the course of maintaining a healthy body, the immune system is far from being circumspect. We have evidence of the potential for an all-out war; “immunological warfare” can be so “blind” that it often also battles its own cells and tissues in a vigorous response to fight off foreign invaders. How do organisms quickly make whole armies of new antibodies to fight entirely new microbial invaders? We can imagine an ancient scene in which a precursor of multicellularity facing such an implacable war with its own limited recourse of somatic embryogenesis (we are assuming that the other modes of development, epigenesis and preformistic, have not evolved yet) had to succumb to or survive the invasive microbial world surrounding it. Natural selection had an urgent task to face, to multiply the new constellation of genes that mutated in favor of pathogen recognition and the regulation of immune responses. 1. Natural selection’s battle starts during the development of the embryo. The first challenge for the immune system would have to be the cells’ own molecular components (self antigens). 2. The first mechanism that should have arisen in the immune response is the one that enables the recognition of microbes. However, this situation presents a fundamental problem for the immune system: to recognize a vast range of foreign pathogens and at the same time to stop short of destroying self. 3. The ontogenetic process had to create by trial and error, thanks to natural selection, the mechanism whereby the interstitial cell multipotent cell lineage becomes something analogous to the immune cell populations of T and B lymphocytes. Strong selective pressures (orthoselection) and time, in favor of alleles that coded for designing interstitial cells made to fight a vast range of foreign pathogens, were presumably the path for an efficient immune system. 4. We propose that a cascade of selected mutants, subjected to the proper stimuli, gave rise to all of the various somatic cell types. Interstitial cells, which in Hydroids can be recognized by their distinctive amoeboid morphology, have been observed to give rise to various somatic lineages, and therefore, could be candidates to harbor mutants destined to initiate lymphocytes. Several researchers (according to Buss, 1987) are actively testing the possibility that the morphological identity of interstitial cells masks an underlying complex of partially differentiated stem cells (lymphocytes). 5. If the above hypothesis turns out to be correct we may have, at this early stage in the evolution of the immune system, the first category of somatic cells, continuously replenished by the differentiation of interstitial cells. 6. The ameboid morphology of differentiated interstitial cells permits the free movement necessary to encounter invading pathogens. GENE SELECTION OR HIERARCHICAL SELECTION Both the gene selectionist view and the hierarchical view are useful for identifying and describing the evolutionary innovations of the somatic immunological phenomena in the first multicellular organisms. To adopt Williams’ (1996) and Dawkins’ (1976) reductionistic interpretation of genes as the only units of selection, and the other candidates for potential units of selection as merely “vehicles” that transport immortal genes from generation to generation, may not be altogether wrong; it simply does not help to unravel and to identify how evolutionary innovations came about. The most difficult problem is identifying and describing how a particular selfish innovation has channeled subsequent evolution. The sole gene selectionist perspective offers no particular guidance in this respect. The hierarchical view recognizes that both the cell and the individual are viable units of selection. In other papers we analyzed whether the different levels of the biological hierarchy - genes, chromosomes, cells, species and even communities - constitute different survival entities that require different heritable variations in fitness in order to function as units of selection in the evolutionary process (Hoenigsberg, 2002). For the theory on the origin of the immune system in the first metazoa that we are now offering, we accept the idea that the above mentioned entities retained to varying degrees some of the primordial lower-level identity, where selfish genes were instrumental to better multiply their type. This lower-level unit is the ideal milieu for natural selection’s survival of the fittest to dominate evolutionary trends. However, other higher levels became necessary in order to channel evolution in the direction of multicellularity. The evolutionary process went through transitions in the units of selection, from protogenes to gene networks and to the various biological steps in the hierarchy of life by inventing cooperation, through opportunistic conflict modifiers and synergic interventions among the lower level units in the functioning of the higher-level unit. One of the greatest evolutionary breakthroughs was the invention of cooperation to regulate the primordial paradigmatic conflictual and selfish program. To understand this momentous crisis, Eigen and Schuster (1979) proposed the hypercycle. In the transition from the small replicators to the cooperating gene groups, as the self-replicating molecule increases in size, it will necessarily suffer a corresponding increase in the frequency of errors during self-replication. These errors would have left larger replicators at a disadvantage compared to the smaller ones. And yet evolution did not stop at small replicants. Eigen and Schuster’s solution is the “hypercycle”, whereby independent replicators, whose autocatalytic properties complemented one another, became attached to one another in self-replicating complexes. Here we have the first case of transition from a lower-level unit of selection to a new higher level. Thus, we are tempted to say that the major features of evolution were shaped during periods of transition between units of selection. The major features of evolution probably include, among many others, the organization of replicating molecules, the metabolic machinery of cells and the principal developmental ‘bauplans’. Undoubtedly, the transition occurring sometime in the Precambrian from the single-cell organisms to multicellular individuals, with division of labor, constitutes one of those innovations. The prior state, that of the free-living totipotent cells, and the subsequent state, that of individuals with differentiated cells, meet the prerequisite for evolution: replication, with occasionally imperfect heredity, producing variants from which selection can distinguish differences. In the theory we are developing here, the transition was marked by a shift in how selection operated on the lower unit. In the prior state, as free-swimming organisms, cells were selected only by the external environment. In the next state, cells were selected according to how they performed in the somatic environment; some escaped terminal differentiation to produce new individuals, others followed cell lineages that conducted them to terminal differentiation. Some cells could not “forget” their previous free-living state and proceeded as before, with ameboid motility maintained together with other new functions, such as rejecting other microbial invasions, cooperating with a population of cells with Germinal-Center functions. The potential for full-scale immunological warfare soon became a vigorous response with strong Darwinian cellular selection in its favor. THE BEGINNING OF THE IMMUNE SYSTEM To start out with the somatic embryogenesis mode of development, the embryo and the earliest mechanism to deal with the relentless barrage of potentially invasive pathogenic microbes had to begin with a powerful promoter of phagocytosis, similar to what we know of today. We think that the first family of molecules were the antibodies, coded by the same housekeeping genes that dealt with the large range of cellular functions already indispensable for unicellular metabolic survival and maintenance (for example the enzymes needed to replicate the DNA, the enzymes and RNA molecules necessary to assemble the ribosomes and allow their functions, the enzymes needed to splice out introns in pre-messenger RNA molecules, etc.), because these are genes with protein/RNA products essential for the basic functions of the cell. Although the first genetic innovation had emerged at the unicellular level as a primordial unit of selection, once the embryo’s cell replication channeled somatic embryogenesis to terminal tissue or cell-specific functions, specific genes mutated out and selected for second order selection, namely the group of cells carrying out a particular specialized function. For example, to deal with “immunological warfare”, involving battles within cells, cell lineages and tissues. We do not intend at this point to deal with the extraordinary ‘intelligence’ of the vertebrate immune system; however, we are proposing that even at this early stage in the evolution of the immune system of metazoa, the first protein antibodies had to exist, such as the pentameric IgM (ten HL heterodimer units) and other antibody proteins, generally abbreviated Ig. The cells that directly derive from the original zygote are clones of it and so we can surmise that the housekeeping genes present in the zygote already started with the variable immunoglobulin genes, which at this time had tissue-specific functions in metazoa, and therefore can be considered to be tissue-specific genes, typically expressed by those cells that should be considered the precursors of B lymphocytes. The same can be said of alpha and beta globulin genes, which were expressed in that lineage of cells, and which later on became red blood cells expressing hemoglobin. In this first immunological challenge there was only a phagocytic response, along with the indispensable protein antibody coat, to make engulfment of viral and bacterial particles possible. Since the five classes of immunoglobulin, termed IgM, IgG, IgD, IgA, and IgE, which have different amino acid sequence proteins in the C-regions of their H chains, display different immunological protective mechanisms, it may not be amiss to suggest that they were all present at that second level of selection, with cellular cooperation (Figures 1 and 2).
Figure 1. Basic structure of an IgG antibody from the modern vertebrate immune system to emphasize the difference from a more simple system that presumably originated from an early metazoan, in which we propose somatic embryogenetic development as a first step in the evolution of the immune response. In the initial development, a distinct germline is lacking. Without a definite germline the successful repulsion of pathogens was subordinated to antigen-driven somatic adaptability. It is possible that antibodies existed in this early beginning but not as complex heterodimers with each basic subunit consisting of two different protein chains as we find in vertebrates today. It seems more likely that such proteins did not have a differentiated variable region (V), a distinct border of the V and C regions and a constant region (C). Moreover, the simple structure of these first antibodies included just one kind of antigen-binding site, instead of the various different kinds presented in this drawing. It is likely that the H and L chains that today interact in the variable regions did not do so in organisms with just somatic embryogenesis.
Figure 2. Diagram showing the complex protein structure of the “intelligent” vertebrate immune system, with many antigen-binding loci. Each antigen-binding site may be of low affinity, but many binding points, as shown in the drawing, increase the overall binding avidity of the antibody. In the advanced metazoan immune system, which evolved an epigenetic mode of development, the immunoglobulin genes (Ig), which are responsible for encoding antibodies that are transmitted to progeny via germ cells and for their preservation throughout evolution, are guided by the same Darwinian laws which apply to other genes. However, our theory emphasizes that in the next unit of selection, where new variable somatically mutated (V) genes are indispensable for individuality to assert its triumph over the “selfish” remnants of DNA sequences; retrogenetic feedback from stem cell lymphocyte mRNA is necessary to further evolutionary development within the germline V gene repertoire. See the literature in Figure 5. HL = heavy and light heterodimer units. All of these antibodies have a membrane-bound form, whereby they are displayed on the surface membrane of the B lymphocytic precursor. IgD, from the very beginning, has had a very important function to perform, which is in the antigen-triggering mechanism, and it is never secreted into the surrounding fluid. IgM, IgG, IgA, and IgE are all secreted from a B cell and I would expect them all to be there in the early evolution of somatic embryogenesis. Possibly IgG, which consists of two heavy and light (HL) heterodimer units, is secreted later in the immune response, and has typical ‘memory’ responses but with very high affinity, important for binding and neutralization of viruses and microbial toxins, could have had just that kind of primary function, although not the more advanced one observed in the vertebrates, where the IgG antibodies produced by the mother cross the placenta and enter the circulation of the fetus. However, IgA, with four HL heterodimer units, could have been present at the very beginning of metazoan evolution as part of the proposed evolvability of this and subsequent organisms that derived their immune system from these early developments, because these proteins served to protect the mucosal surfaces of the digestive tract. IgE, with two HL heterodimer units, responsible for acute allergies and various tissue-specific signals, could have initiated its carrier later on during the emergence of highly complex synergic mechanism between the next two units of selection. I mean, of course, for allergic signaling cooperation to arise, they would have to abandon other tissue-specific functions. It is important to comprehend that if we want to get animals out of the Precambrian developmental chaos (see Buss, 1987; Bonner, 1988) into functional bauplans, we have to think of an organism with germline sequestration, even if that obliges us to admit a particular form of synergism between the lower selfish adaptation of unicellular organisms and the higher individual metazoan multicellular one. This is possible when one adopts the hierarchical view of selection, as we assumed previously. We would not expect the first trial mutants (which emerge from a Germinal Center) from the genome of the B-like lymphocytic precursor to show appropriate affinity maturation of the mutant antibodies, to have higher binding affinity for the antigen than is need at this early phase of the immune response. Although, the immune response had to be quick and definite, it did not need to create the whole intricate three-dimensional folding of the variable regions of an H and L chain to constitute the surfaces of the heterodimer which bind the chemical moieties of the incoming pathogen. The antigen-combining site went through a series of steps until the instrument was perfected for the first so and so adaptation. In the mean time, phagocytosis was enough to free the somatic lineage of pathogens, allowing it to fulfill its terminal tissue function, granting only a ‘certain’ amount of survival and apoptosis. The various classes of immunoglobulin proteins did the old job of defense they had been doing for unicellular haploid organisms, while conflicts between the initial lower-level selection and the higher-cooperative level were at its best. The individual multicellular proposal was a division-of-labor breakthrough, a true innovation that finally prevailed and the second higher-level selection got in to favor cooperation and individuality thanks to diploidy in zygotes. Diploidy was an early invention to suppress the deleterious consequences of selfishness in haploids. At the same time another breakthrough, the eukaryotic nucleus, appeared to subdue conflicting genes with proper modifiers in organized chromosomes (Maynard Smith and Szathmary, 1995). THE GERMINAL CENTER Another early invention in somatic embryogenetic development without a distinct germline was to evolve into a preformistic mode of development. Thus, Weismann (at the end of the 19th century) was correct in concluding that heritability is controlled by development. Another victory of individual selection over gene level selection was sequestering a specific cell lineage from the zygote in the innovative preformistic metazoan development. With this jump into a new evolvability, individuals acquired gamete formation, limited totipotency and immortality. In sexual metazoans, this opened the way to evolvability, of a special area of tissue that, although it did not have all the later functions of the spleen or of the lymph nodes, at least had increased cell division to fight the constant war against the multitude of pathogens. That was the Germinal Center. There was a need for an area where antigen-activated B-like cells undergo intensive proliferation, with rapid divisions every 5-8 h in contrast to cell division times of some 24 h for most other somatic cells. We propose that the original Germinal Center consisted of up to several hundred progeny cells (today’s vertebrate Germinal Centers consist of at least 20,000 progeny B cells derived from one founder B cell). Then, as today, the process of somatic hypermutation was activated in the special cellular microenvironment called the Germinal Centers. THE SOMATIC HYPERMUTATION The mutation process activated by antigenic stimulation of a B cell evolved from a system in which the invading virus or bacteria secreted surface mutagens as a means to counter somatic cell reactions. The flush of mutagens from the pathogens in turn causes today, as it did in the past, the rearranged variable region genes (V(D)J) of the antibody expressed in that precursor of today’s B cell, to mutate the bases in their DNA sequence at a rate a million times faster than the normal background mutation rate. The process of somatic configuration must have originated through Darwinian selection in the rearranged variable region genes represented by the generic symbol V(D)J and found only in mature lymphocytes. Was there a selectively created mutation process in the immune system able to generate new somatic mutants of Ig antibody genes in response to persistent (generation after generation) massive microbial invasions? Was this still another strategy for the individual selection to triumph over the ‘local selfish Ig genes’ that were responsible for proliferating themselves at the expense of the health of the individual? The mechanism was indeed very appropriate and simple, as it produced pre-existing antibody specificity variants before foreign antigens impacted on the system. After all, why should the immune system generate new somatic mutants of antibody genes in response to a foreign antigen? In the evolution of the early immune system, when somatic lineages were still totipotent stem cells, the few Ig genes needed to propagate antibody defenses in the germline DNA were the starting family of genes that later responded to the more invasive entrance of viral particles in animals with backbones, by selectively maturing stems cells that effectively accumulated mutants in the somatic environment to confront those invasions that needed an immediate effective response. It is our view that from then on individual selection set up a double-immune system, with germline DNA to take care of propagating to future generations the defense capabilities, and a second system, arising during the lifetime of the terrestrial animal by somatic mutation stimulated by antigen (Figure 3).
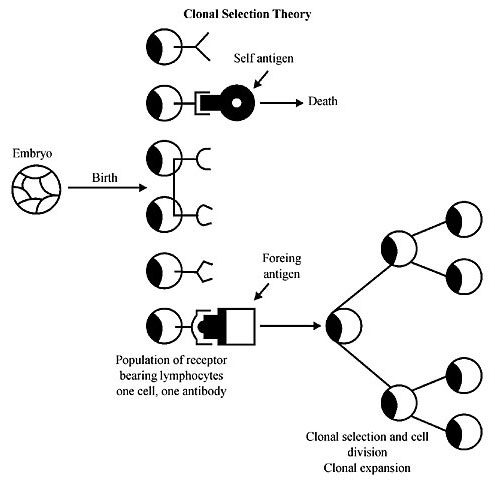
Figure 3. A very simple diagram for the Clonal Selection Theory, developed by MacFarlane Burnet to solve self versus non-self discrimination problems. For Burnet the cell is the basic unit selected by an antigen, and that one cell is responsible for producing one type of antibody. This extraordinary idea developed the central theme that ‘one cell makes one antibody’ and that many different cells (lymphocytes) exist and each one (in this new metazoan invention, in our theory, a new triumph over the gene as the unit of selection), when Darwinianly selected by the antigen which fits the shape of the antibody, multiplies to give a clone of identical cells all producing the same antibody. The ‘decision’ to express one antibody per B cell takes place during early maturation of the lymphocyte in today’s vertebrate immune system. However, in early metazoan evolution (before their epigenetic mode of development) when somatic functions of stem-cell-lymphocytes were just beginning in the cellular environment of the Germinal Center there were somatic cell selection ‘decisions’ that determined efficacious control of viral invasions. It was that early cellular unit of selection that defined the course to take in the evolution of the immune system through antigen-binding selection. THE SEPARATION OF THE GERMLINE CONFIGURATION FROM THE SOMATIC ONE TO OBTAIN TWO DIFFERENT SYSTEMS RESPONDING TO TWO DIFFERENT UNITS OF SELECTION The immune system of ancient metazoa was challenged by a relentless barrage of potentially invasive pathogenic viruses and bacteria. It is possible that the somatic lineages did not have the synergistic relationships between the cellular and the genetic unit of selection needed to resolve the problem from that angle. It was necessary to wait for individuality to emerge as a process to regulate a unit of selection besides cellular proliferation and competition. The cells of a metazoan are endowed with the capacity for heritable variation and differential replication; in other words, all the prerequisites for evolution. Heritability and the variation that results are not denied to the cell lineages within an organism, nor are the cell lineages outside of the rules that govern any self-replicating system. Thus, variants that enhance the replication rate are favored. Perhaps we should insist in this essay that variations at the level of the cell lineage may be inherited by lineal descendants within the organism; however, they are not passed on to subsequent generations. There are consequences for a cell lineage that becomes terminally committed to some somatic duty; it is precluded from ever giving rise to gametes. At the level of the individual, heritability is precluded by terminal somatic differentiation. While the cell lineage may not be able to produce sex cells, heritability at the level of the cell lineage is not excluded. Those factors that increase the replication rate of cell lineages within the organism may be deleterious to the individual as a functioning unit, although they will be strongly favored at the level of the cell. Thus, to produce a harmoniously functioning unit (such as a terrestrial vertebrate immune system) mechanisms must have evolved whereby variants that selfishly enhance their own replication rate by not adopting somatic duties are controlled. Modifiers of conflict by controlling cell lineages, conflicting genes and mutants that deviate from their somatic duties must evolve in the evolution of the system. Selection at the level of the individual must have effectively opposed selection at the level of the gene of the cell lineage. The metazoan embryo is not excused from this conflict. The ontogeny of a metazoan involves a continued sequence in which cell lineages progressively deny their own capacity for increase to favor individuality. The conflict between the potentially opposing processes of somatic differentiation and individual harmony has been resolved in favor of the individual. The existence of a harmoniously functioning vertebrate immune system in multicellular organisms is compelling testimony of this fact. Metazoan evolution is characterized by an increasing sophistication of cells, groups or populations of cells, tissues and organs that perform somatic jobs indispensable to the organism as a whole, but which require for the cells composing them to limit their inherent capacity for proliferation. However, the genetic program for continued self-replication momentarily subjugated to the interest of the whole lurks behind the scene in order to escape conflict modifiers (Figure 4). In what moment of the development of the embryo, or rather how did the maternal mRNA instruct embryonic development to start synergism between the cellular and the individual unit of selection to develop a somatic configuration separate from a germline configuration?
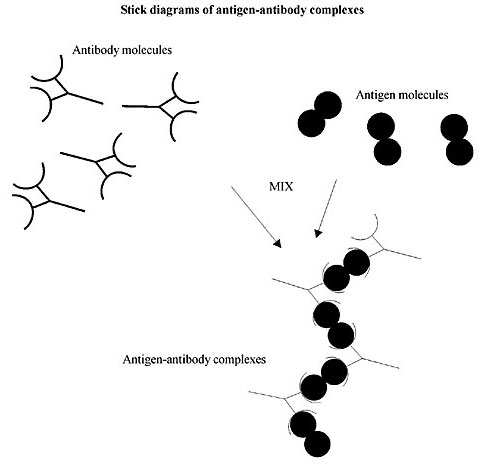
Figure 4. In our theory, the early metazoan with a somatic embryogenetic mode of development had one cell lineage capable of participating in detecting and refusing microbial infection with a very specific somatic function as a stem cell (precursor of lymphocytes). But it is also capable of accomplishing germline functions throughout ontogeny (as in plants). During this primordial day “selfish” developments were still in vogue notwithstanding that cooperation had initiated individuality. But the “selfish” interests of genomic structure, cellular organization (in somatic proliferation), and ontogenetic patterns manifestly produce “vehicles” (alla Dawkins) in the form of organelles, cell populations, etc. Those selfish pursuits that favor the replication of both the gene and the vehicle are more Darwinianly likely to survive than those that favor the gene, but not the vehicle. Germinal Centers are vehicles with intense cell proliferation and cell death (apoptosis) of lymphocytes caused by antigenic stimulation. In this diagram, antigen-antibody complexes bind to the surface membrane of cells such as phagocytes via the constant segments of the antibody H chains, enabling their engulfment and digestion. Synergisms between genes and vehicles, in times when the individual unit of selection is still incomplete, predict that Germinal Centers’ special cells called the follicular dendritic cells are vehicles for the selection of high-affinity variable region mutants. THE CELL EXPRESSES THE ANTIBODY ON ITS SURFACE WHILE ITS MATURATION PROCEEDS The genes encoding V- and C-domains are physically separated by a long stretch of DNA (V is some 300,000 bases upstream of the C-portion!) for an (as yet) unknown reason. Ig genes are responsible for both the H (heavy) and L (light) antibody chains. They code for an amino acid sequence called the V-domain (for antigen binding) and a constant or C-domain (e.g., for triggering target cell lysis or phagocytosis), this could be an ancient constant DNA stretch. While the housekeeping genes and the other regular genes are located in specified positions on the chromosomes, antibody genes present a very special situation. The genes for the V- and C-domains of heavy chains in the mouse are on chromosome number 12, which is towards the end of the long arm, near the telomere. Yet genetic mapping and DNA sequencing show that the V-portion is at least 300 kb upstream of the C-portion. How can this be? Many years ago Dreyer and Bennett predicted that the contiguous stretch of DNA encoding an H- and L-protein chain is produced through a chromosomal translocation when the V-portion of the DNA rearranges its sequence position to come close to the C-portion. That means that the question ‘how can that be?’ could be solved by speculating that there was splicing during the maturation process of lymphocyte cells. The genius of the speculation made by Dreyer and Bennett was even greater when they suggested not only that the upstream portion of the Ig locus encoded a large number of ‘germline’ V genes in any one single B cell, but also that the rearrangement consisted of shifting one of these V genes to a place close to one of the possible C-region genes (see the account given by Steele et al., 1998). This could be a strategy to increase randomness in the chosen specificity of an antibody. By drawing just one V gene at random from a large number of them, cellular selection would enhance its probability of getting the desired unique specificity. This random rearrangement increases the number of unique surface-type antibodies capable of fitting, allowing the binding of an enormous number of different antigens thanks to the great ensemble of complete antibody proteins of previously unknown specificity from a large number of possible combinations of the H- + L-protein chains. If the combination happens to be anti-self it would be deleted following the self-tolerance of the forbidden clones of Burnet’s clonal selection theory. In fact Tonegawa, who revealed the organization and rearrangement of genes in the Ig loci of mice, proved the initial bold prediction of Dreyer and Bennett (Figure 6 describes just that). When the separate elements of antibody genes in the non-rearranged sequence appear, the format of the DNA is called the ‘germline configuration’ (which is the same sequence found in DNA from germ cells and in all the other cells of the body that are not mature lymphocytes). On the other hand, the ‘somatic configuration’ format is the name given to the rearrangements of the heavy chains in each B lymphocyte, during B cell development in the bone marrow, in V, D and J elements, so as to produce a rearranged variable region sequence, VDJ. First of all, the H chains rearrange the fused DJ sequences randomly and secondly another random fusion with an upstream V element makes a VDJ. Thus, each successful variable region rearrangement is special and unique to each maturing B cell. The pre m-RNA containing the antibody variable region (VDJ) attached to the C-region is made in the nucleus. What results after splicing of the L-V introns and the J-C introns plus the introns of the C-region in the pre-mRNA sent out into the cytoplasm is the mature mRNA. After that maneuver, translation in ribosomes produces the desired and unique H chain, followed by the constant region and the variable piece with the leader sequence clipped off (see Figure 6). This amazing stunt is no doubt the making of a selection mechanism that evolved through a series of trials that resulted in debilitated individuals at first, until it could produce the large number of diversified surface antibodies needed to attend most microbial infections. In the germline configuration the DNA cannot be transcribed into mRNA, so that antibodies cannot be synthesized. In the somatic configuration rearranged V(D)J and C genes are actively transcribed, resulting in the production of mRNA, which is then translated into H and L chains to form an antibody. The somatic mutation process is only active in the V(D)J rearrangements and never in the unrearranged V genes in the germline configuration. As usual, the rearrangement of this ‘acrobatic feat’ is executed by a set of recombination enzymes encoded somewhere else in the genome by the now famous recombination activating (RAG) genes, which themselves must be instrumental in this surgical operation. I will come to that later. As of now we have it clear enough that the whole operation of leaving one region to mutate, the V-region, while preserving the C-region intact made it possible for individual selection to build a process capable of a rapid and effective Lamarckian feedback loop between the soma and the germ cells; this was accomplished with Darwinian selection first and the Lamarckian procedure after. This, in the long history of transitions in evolution, has been the final triumph of the individual over the selfish gene (Figure 5).
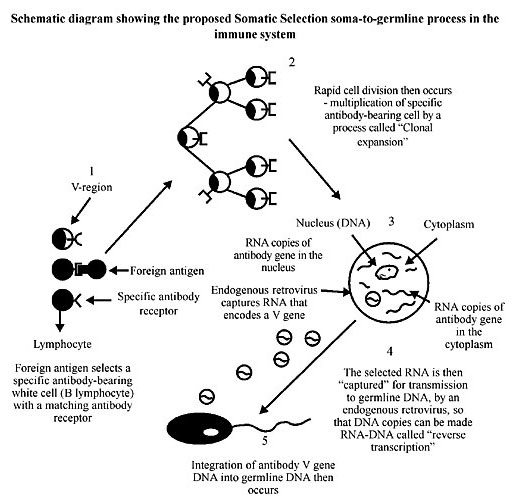
Figure 5. Schematic diagram showing the proposed ‘Somatic Selection’ soma-to-germline process in the immune system proposed by Steele (1979) and modified from Steele et al. (1998), and to which we adhere for late metazoans with epigenetic development that possess a clearly differentiated germline. Such organisms developed a germline after the primordia of major organ systems of the adult had become established (e.g., annelids). Our adherence to this Neo-Lamarckian theory extends even to the other extreme of the embryological timing that specifically includes the terminal differentiation of the germline, in organisms with preformistic development (e.g., mammals, birds, mice and humans). Here the germline is terminally differentiated in earliest ontogeny, in which orientation of maternal-derived determinants deposited in the egg is present. 1. A diverse array of B lymphocytes exists before the entrance of a foreign antigen into the system. Each cell exposes on its surface membrane antibodies of one unique specificity. The variable (V)-region genes of the lymphocytes encode those parts of the antibody that form the antigen-combining site. A foreign antigen binds to those B cells which have compatible antibodies; therefore, these cells are ‘selected’ in a Darwinian manner, generally called clonal selection. 2. After antigen binding the B cell is activated and divides, producing progeny profusely that in turn divide, producing more identical progeny cells. A clone of identical cells expressing that antibody is produced (this is called clonal expansion). Individual cells of the clone can mutate their variable region genes (see Figures 2 and 6) that can themselves be selected by an antigen to produce another clone. 3. Within cells of the clone, RNA copies of antibody V genes are made in the nucleus. Mature mRNA is sent to the cytoplasm where it is translated into the appropriate sequences of amino acids making up the protein chains of the antibody. 4. RNA molecules encoding V genes (in the nucleus of the cytoplasm) may be engulfed by innocuous endogenous RNA retroviruses (produced by the cell) and complementary DNA copies of the RNA made by the viral enzyme, reverse transcriptase. These are the molecules that probably belong to the ancient RNA world and which were discovered by Temin in 1959 (see Temin, 1989) and by Temin and Baltimore in RNA tumor viruses in 1970. 5. According to Steele (1979) and to Steele et al. (1998) Neo-Lamarckian theory, which we adhere to for late Metazoan epigenetic organisms, and to the other extreme of the ontogenetic timing, for those with a preformistic mode of development, copies of the antibody V genes are transferred (alla Temin and Baltimore) by the virus with the inserted somatically mutated characteristic to sperm or egg nuclei. Thus, the germline DNA is enriched with more essential elements of the antibody repertoire. See a molecular unifying hypothesis (Blanden and Steele, 1998). MUTATIONS CONFINED TO V(D)J REGIONS IN THE SOMA-TO-GERMLINE FEEDBACK IN STEELE’S SOMATIC SELECTION AND ADAPTIVE EVOLUTION From the last paragraph it is evident that while mutations in the variable portions of antibodies are very useful, mutations in the constant portions can be dangerous to the system because they can render it useless for triggering the lysis of microbial cells or for promoting phagocytosis. Thus, separating the V- from the C-region genes through a quick cytological process has allowed the evolution of mechanisms that selectively mutate the V-region while preserving the C-region. This is what happens during the maturation of B lymphocytes, and high rates of somatic mutations at the V(D)J gene and the resulting antibodies can be Darwinianly selected for utility in antigen binding. The coefficient of selection gives the prize to those with the highest affinity for the antigen, as the lymphocytes producing them are retained to become long-lived memory cells. Obviously, those mutations that reduce or cancel antigenic affinity cause the B lymphocyte to lose the prize and die in the classical Darwinian manner. Those B cells encoding antibodies that bind to the self-antigens are eliminated by a special process, thus preserving self-tolerance. We cannot explain all the mechanics involved in the estimates of the size of the functional antibody repertoire. At this point we can suggest Langman and Cohn with their Protection Theory (1987) or the V Gene Replacement Theory proposed by the group led by Weigert (Chen et al., 1995). What is relevant here is that now we know the molecular details of the separation of the germline configuration from the somatic configuration. With our interpretation of the molecular data and the theoretical interpretation of the possible embryological and evolutionary transitions in terms consistent with a Lamarckian soma-to-germline recourse we are in a position to answer the question of how the embryological and the synergistic action between the gene-cellular unit of selection suggested in the Clonal Selection theory of Burnet in 1957 and the individual unit of selection could have operated together to increase the variable genetic repertoire of the vertebrate immune system. The unique features of Ig genes, and the recognition that the process of developmental control heritability transfer the focus of the issues of inheritance from the purely molecular mechanisms of transmission to questions of patterns in developmental determinism. Thus, the ancient metazoan embryological epigenetic mode of development with a clearly differentiated germline permits a new interpretative framework for understanding the new genetics of antigen-recognition molecules. This section of our essay is based on the known RNA splicing patterns in V(D)J messenger RNA, reverse transcription (copying of RNA back into DNA) and the shuttle to take the somatic DNA to the germline (Figures 6 and 7).
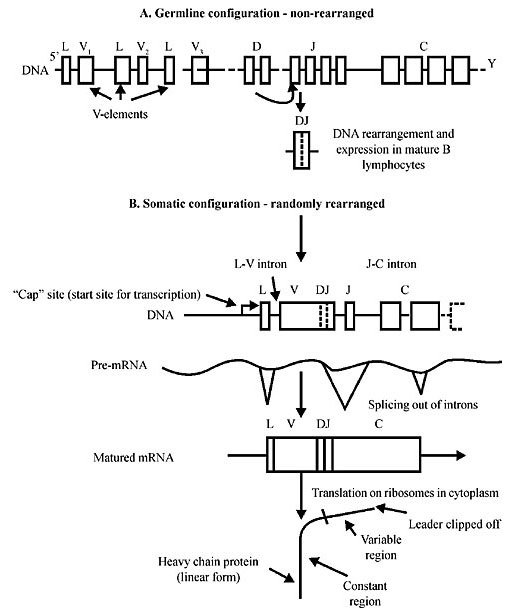
Figure 6. Schematic outline of a) the germline DNA sequences in the upstream variable gene (V) elements of the constant (C) region-coding exons. Such germline V-genes are ‘non-rearranged’. In the drawing, between the V and C are the diversity (D) and joining (J) region elements. At the end of the V-elements stretch there is a leader sequence (L) which encodes a short sequence of amino acids, referred to as a ‘signal peptide’. All genes-encoding proteins, such as antibodies destined to be sent from the cell, or to another membrane-bounded compartment within the cell, have such ‘signal peptides’, which allow the system to recognize them and be shunted to their correct cellular or extracellular locations. b) During B cell development (this applies to the advanced vertebrate immune system) in the bone marrow, each lymphocyte randomly rearranges its heavy chain V-, D- and J-elements so as to give a rearranged variable region sequence (in the mature B lymphocyte) VDJ, now said to be in the ‘somatic configuration’. In order to appreciate the extraordinary uniqueness of each maturing B cell in the vertebrate immune system, we describe in detail how the intervening DNA between the V-element chosen by chance for rearrangement and the J-gene sequence utilized are looped-out and deleted from the cell. In fact the first step in the rearrangement is the random formation of a fused DJ, followed by the random fusion of an upstream V-element to create a VDJ. There is also the splicing of the introns and the resulting mature mRNA. After translation on ribosomes the protein H chain of the antibody has its C- and V-regions and the Leader region is clipped off. We have used Steele et al. (p. 113, 1998) liberally, but with some modifications.
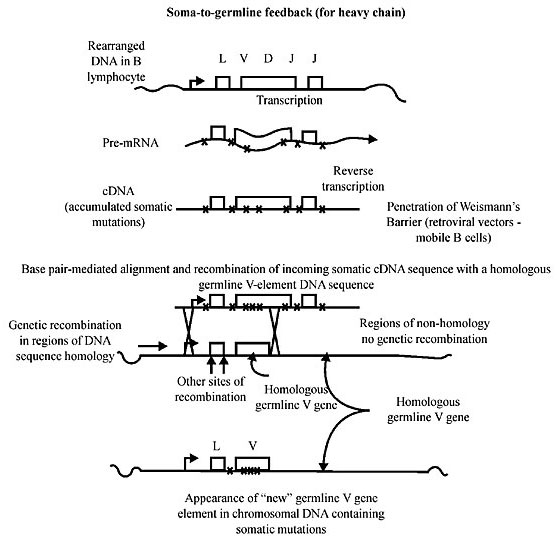
Figure 7. The outline shows the end result of the Neo-Lamarckian soma-to-germline feedback to which we adhere and which should be understood to represent an extreme form of synergism between the gene level of selection and the individual level, where the latter proclaims its final triumph in the vertebrate immune system (Blanden and Steele, 1998). As explained before (see the literature in Figure. 2) somatic mutations accumulate in the variable V(D)J genes of a B lymphocyte (as discussed in Figure 6). Mutated reversed transcripts (cDNA) accumulate as somatic mutations and penetrate Weismann’s Barrier with retroviral vectors found in the cytoplasm of the lymphocyte and are delivered to germ cells, where they integrate into germline DNA by the well-known process of ‘homologous recombination’. In the drawing there are other sites of potential recombination but also other regions in the chromosome of the germline of non-homology and therefore of no recombination. In Steele et al. (1998) the reference to the work of Weiller et al. (1998) is made to explain how the recombination profile of germline V genes was established by use of the Philopro genetic recombination algorithm. It appears that the major sites of the germline recombination algorithm are at the transcription start site, the borders of the L-V intron and the end of the V-coding region...this constitutes the discussed ‘integration footprint’. THE SOMATIC MUTATION For a review of the original scientific literature about the somatic mutation in the vertebrate immune system, see Steele et al., 1993, 1997, 1998. For an argument that, from an evolutionary point of view, somatic hypermutation and affinity maturation could be redundant, see Zinkernagel group’s results (Roost et al., 1995; Bachmann et al., 1997). For brilliant and imaginative experiments concerning this topic, see Neuberger and Milstein (1995). And for a review on the somatic hypermutation field prior to Neuberger and Milstein’s paper, see Steele (1991). The transition between the gene as a unit of selection and the cellular level emerges when the mechanisms of previously random and deleterious mutations are subdued for the benefit of individuality. Selection had to avoid overdoing the mutational change and its randomness. Synergism at the two levels of selection had to produce a system in which random errors in the DNA sequence were left out and only those mutated genes that successfully meet the selection criterion were retained. There was still another critical point to resolve, namely that successful mutated sequences do not change to unsuccessful ones. It appears that the solution was to remove the successful ones from the Germinal Center and into a shuttle that can take them to the germline; in other words, into a retrovirus after the proper treatment changed RNA into cDNA. The passage from an adaptive mutation transcribed into mRNA to the cDNA, through reverse transcriptase, had to be rapid to avoid new mutations. Clearly not all gene mutations are harmful. There are those of a special class that confer an advantage to the immune system of vertebrates. As we have seen in the scientific literature, there is considerable evidence that points to mutations in the V(D)J genes expressed in mature lymphocytes and that later appear in the high affinity antibodies (these are highly adaptive and they fit better to foreign antigens). Such antibodies are characteristic of long-lived memory B lymphocytes. There is considerable molecular evidence documenting the fact that high-rate antigen-driven somatic mutations targeted to the variable antibody region V(D)J genes exist. FINALE The late 19th century biologist, August Weismann, created the modern biological definition of the individual. Today we see individuality as a final product of a hierarchical view based on multiple units of selection. Indeed the greatest of August Weismann’s legacy is his “Doctrine of the Continuity of the Germ Plasm”, and with it the view of individuality as seen in modern biological theory. Weismann’s great contribution was his discovery that heritability, and therefore evolution, is controlled by development (for a recent treatment, see Hoenigsberg, 2002). Since terminal somatic differentiation denies a cell lineage the opportunity to contribute to subsequent generations, evolution repaired that synergistic emergency between the gene, the cell and the individual units of selection, by using, in the final triumph of individuality, i.e., the vertebrate ‘intelligent’ immune system, the Neo-Lamarckian selective mechanism, and the Somatic-Germline-Feedback. Plants, colonial invertebrates, fungi and many other organisms with different modes of development violate Weismann’s doctrine, displaying enormous phenotypic plasticity and considerable ecophenotypic variation. In the heydays of Modern Synthesis, for genetics to develop it was necessary to establish that the acquired characteristics could not be inherited, notwithstanding that Weismann’s barrier was frequently found violated and that many important experimental results in Protozoa (see the inheritance of antigenic characteristics in Paramecium) in several works of Sonneborn (1930, 1964) or the works of Waddington (1953, 1957, 1961), which he preferred to call genetic assimilation, indicated the transmission of some environmental insults (see Sapp, 1987). There were also theoretical papers and reviews of experimental contributions, like the one by Zuckerkandl and Pauling (1962) and Zuckerhandl, 1986 on the Dormant Hypothesis, or the Steady State System, preceding the self-replicating nucleic acid, implying environmental transmission and transmigration by Dyson (1985), or the Theory of the Origin of Life by Cairns-Smith (1985), based on the self-assembly and reactions of complex organic molecules, meaning that the epigenetic inheritance system (EIS) probably also had a very early origin that could have preceded the emergence of an inheritance system based on nucleic acids. We should not discard self-sustaining systems that some have called steady-state EIS (Jablonka and Lamb, 1999). We have insisted that development has oriented and determined inheritance and that having left it out of the theory of evolution has been an error. The molecular biology of EIS can broaden Darwinian theory and possibly can give an explanation of some aspects of evolutionary history that are more satisfying than that based on natural selection of random DNA variations. The presence of a mosaic of isochores (Bernardi, 1989, 1993 and many more publications by his group) in the genome of vertebrates has received a very interesting evolutionary explanation in that they have evolved through their effect on thermal stability. GC-rich DNA is more stable than AT-rich DNA at the body temperature of warm blooded vertebrates. It seems that many of the codons contain G and C code for amino acids that contribute to the thermostability of proteins. Therefore, it follows that natural selection in a perfect Darwinian process has favored AT to GC changes in the genes of functionally important proteins. Nonrandom distribution of long interspersed repeated elements (LINE) or short interspersed repeated elements (SINE) is the result of a need for retrotransposons to accommodate them (Okada, 1991; Martin, 1991). For example, an AT-rich transposon will be found in AT-rich regions because it ‘prefers’ this region (the region is more ‘hospitable’ as it were to such a transposon). Just to show that it can be a matter of finding a nonconventional interpretation, Jablonka and Lamb (1999) have offered another angle: they say that rather than being tolerated or not by the regions into which they insert LINE, they have been important in bringing about stable repression of genes in the dark G bands. In other words, LINE could be sequences that facilitate the binding of proteins that repress gene activity (Martin, 1991). By so doing long-term repression may be easier when genes act in blocks rather than as single units. Thus, Jablonka and Lamb suggest that a need for stability and long-term epigenetic memory has determined the coding and noncoding DNA organization. Therefore, it is reasonable to state that environmental factors can directly influence DNA sequence divergence through their effects on chromatin structure. In what way does this happen? And when did it happen? Are we confronted with another case of conflicts between two units of selection, a gene as a unit of selection versus the individual unit of selection, as we have suggested for germinal configuration and somatic configuration in the vertebrate immune system? Then the role of the environment in evolution cannot be taken as only an instrument of selection. If Bernardi’s isochores can resist this interpretation, then the environment can be an agent of variation as Sonneborn suspected many years ago. The Modern Synthesis never generated a theory of ontogeny simply because it assumed that individuality was a fundamental assumption for genes to determine evolution in populations by simply changing frequencies. Therefore, the modern synthesis cannot be expected to explain how individuality evolved. Coming back to the conventional nucleic acid world, theoretical biology has to admit that evolution is the history of transitions between different units of selection (Michod, 1997a,b, 1999). In this essay what we have is a series of transitions that probably started with 1) the origin of self-replicating molecules, 2) the fusions (some strong, others loose combinations) of autonomously replicating molecules into self-replicating complexes, 3) the incorporation of such complexes into elemental cells, and 4) the incorporation of many organelles capable of autonomous replication and many more transitions that led to the invention of sexuality and with it the origin of the species. Transitions were the instruments of general selection to solve conflicts between genes, at the first level of selection, between genes and their self-replicating complexes, between these and their consolidated higher units, between the latter and the cells with organelles capable of autonomous replication, etc. Bringing our hierarchical view to the embryo, there is the invention of the germline as a sequestered group of a few cells from the embryo, the stem cells as a new transition to solve conflicts between cells with terminal development, and involvement of the most important characteristic of metazoan development, the constant fight to survive massive invasions of pathogens, that not only debilitated individuals but also constituted the most potent selective mechanism to consolidate individuality. In the vertebrate immune system we have excellent examples of synergism between different units of selection: at the gene, at the cellular and at the level of the individual. Germline sequestration represents a particular form of synergism between the cell and the individual. Both, however, have to be viable units of selection, and both can retain selfish adaptations at both levels of organization. Stephen Jay Gould, among a few others, has emphasized the role of synergism and conflicts in the interpretation of the evolution of certain biological phenomena. He has used sexuality as the model to present his point; we have used the vertebrate immune system as our model (notwithstanding Brent et al., 1981 and 1982 criticism of Steele’s 1979 theory) but we have gone further, because we have used Darwinian Selection to pave the way to New-Lamarckism in order to explain how nature arrived at the triumph of individuality with the extraordinary contrivances to create a most ‘intelligent’ immune system. We have missed many of the chapters of evolution by not merging genetics with development and by taking the modern synthesis as a dogmatic paradigm. REFERENCES Bachmann, M.F., Kalinke, U., Althage, A., Freer, G., Burkhart, C., Roost, H.-P., Aguet, M., Hengartner, H. and Zinkernagel, R.M. (1997). The role of antibody concentration and avidity in antiviral protection. Science 276: 2024-2027. Bernardi, G. (1989). The isochore organization of the human genome. Annu. Rev. Genet. 23: 637-661. Bernardi, G. (1993). The vertebrate genome: isochores and evolution. Mol. Biol. Evol. 10: 186-204. Blackstone, N.W. and Ellison, A.M. (1998). Metazoan development and levels of selection. Unpublished manuscript (cited by Richard E. Michod, Darwinian Dynamics, 1999, Princeton University Press, Princeton, NJ, USA). Blanden, R.V. and Steele, E.J. (1998). A unifying hypothesis for the molecular mechanism of somatic mutation and gene conversion in rearranged immunoglobulin variable genes. Immunol. Cell Biol. 76: 288-293. Bonner, J.T. (1988). The Evolution of Complexity. Princeton University Press, Princeton, NJ, USA. Brent, L., Rayfield, L.S., Chandler, P., Fierz, W., Medawar, P.B. and Simpson, E. (1981). Supposed Lamarckian inheritance of immunological tolerance. Nature 290: 508-512. Brent, L., Chandler, P., Fierz, W., Medawar, P.B., Rayfield, L.S. and Simpson, E. (1982). Further studies on supposed Lamarckian inheritance of immunological tolerance. Nature 295: 242-244. Buss, L.W. (1987). The Evolution of Individuality. Princeton University Press, Princeton, NJ, USA. Cairns-Smith, A.G. (1985). Seven Clues to the Origin of Life. Cambridge University Press, Cambridge, MA, USA. Campbell, R.D. (1983). Identifying Hydra species. In: Hydra: Research Methods (Lenhoff, H.M., ed.). Plenum Publishing Corporation, New York, NY, USA, pp. 19-28. Chen, C., Nagy, Z., Prak, E.I. and Weigert, M. (1995). Immunoglobulin heavy chain gene replacement: A mechanism of receptor editing. Immunology 3: 747-755. Dawkins, R. (1976). The Selfish Gene. Oxford University Press, Oxford, England. Dyson, F.J. (1985). Origins of Life. Cambridge University Press, Cambridge, MA, USA. Eigen, M. and Schuster, P. (1979). The Hypercycle, a Principle of Natural Self-Organization. Springer-Verlag, Berlin, Germany. Ewer, R.F. (1948). A review of the Hydridae and two new species from Natal. Proc. Zool. Soc. Lond. 118: 226-244. Gerhart, J. and Kirschner, M. (1997). Cells, Embryos, and Evolution. Blackwell Scientific Publications, Cambridge, MA, USA. Giordano, R., Magnano, A.R., Zaccagnini, G., Pittoggi, C., Moscufo, N., Lorenzini, R. and Spadafora, C. (2000). Reverse transcriptase activity in mature spermatozoa of mouse. J. Cell Biol. 148: 1107-1113. Godelle, B. and Reboud, X. (1995). Why are organelles uniparentally inherited? Proc. R. Soc. Lond. Ser. B. Biol. Sci. 259: 27-33. Grayson, R.F. (1971). The freshwater Hydra of Europe. I. A review of the European species. Arch. Hydrobiol. 68: 436-449. Hoekstra, R.F. (1990). Evolution of uniparental inheritance of cytoplasmic DNA. In: Organizational Constraints on the Dynamics of Evolution (Maynard Smith, J. and Vida, G., eds.). Manchester University Press, New York, NY, USA, pp. 269-278. Hoenigsberg, H.F. (2002). The future of selection: individuality, the twin legacies of Lamarck & Darwin. Genet. Mol. Res. 1: 39-50. Jablonka, E. and Lamb, M.J. (1999). Epigenetic Inheritance and Evolution. Oxford University Press, Oxford, England. Landman, O.E. (1991). The inheritance of acquired characteristic. Annu. Rev. Genet. 25: 1-20. Langman, R.E. and Cohn, M. (1987). The E-T (Elephant-Tadpole) paradox necessitates the concept of a unit of B-cell fuction: The protection. Mol. Immunol. 24: 675-697. Lewin, R. (1983). How mammalian RNA returns to its genome. Science 219: 1052-1054. Litman, G.W. (1996). Sharks and the origin of vertebrate immunity. Sci. Am. 275: 47-51. Martin, S.L. (1991). LINEs. Curr. Opin. Genet. Dev. 1: 505-508. Maynard Smith, J. and Szathmáry, E. (1995). The Major Transitions in Evolution. W.H. Freeman, San Francisco, CA, USA. Michod, R.E. (1997a). Cooperation and conflict in the evolution of individuality. I. Multi-level selection of the organism. Am. Nat. 149: 607-645. Michod, R.E. (1997b). Evolution of the individual. Am. Nat. 149: 607-645. Michod, R.E. (1999). Darwinian Dynamics. Princeton University Press, Princeton, NJ, USA. Neuberger, M.S. and Milstein, C. (1995). Somatic hypermutation. Curr. Opin. Immunol. 7: 248-254. Okada, N. (1991). SINEs. Curr. Opin. Genet. Dev. 1: 498-504. Pollard, J.W. (1984). “Is Weissmann’s Barrier Absolute?” In: Beyond Neo-Darwinism: Introduction to the New Evolutionary Paradigm (Ho, M.W. and Saunders, P.T., eds.). Academic Press, London, England, pp. 291-315. Prasad, N. and Mookerjee, S. (1986). Morphogenetic analysis of ecotypes of Indian Hydra. Part III. Understanding Physiological responses. Zoological Survey of India. Tech. Mong. 13: 75-108. Ransick, S., Cameron, R.A. and Davidson, E.H. (1996). Postembryonic segregation of the germline in sea urchins in relation to indirect development. Proc. Nat. Acad. Sci. USA 93: 6759-6763. Rastogy, N., Pandey, M. and Mookerjee, S. (1992). Statistical analysis of geographical variability in 16 ecotypes of Indian Hydra. Evol. Biol. 6: 195-204. Roost, H.P., Bachmann, M.F., Haag, A., Kalinke, U., Pliska, V., Hengartner, H. and Zinkernagel, R.M. (1995). Early high-affinity neutralizing anti-viral IgG responses without further improvements of affinity. Proc. Nat. Acad. Sci., USA 92: 1257-1261. Rothenfluh, H.S. and Steele, E.J. (1993). “Origin and Maintenance of Germ-line V-genes”. Immunol. Cell Biol. 71: 227-232. Rothenfluh, H.S. and Steele, T. (1993). “Lamarck, Darwin and the Immune System”. Today’s Life Sci. 5: 8-15 and 5: 16-22. Rothenfluh, H.S., Blanden, R.V. and Steele, E.J. (1995). ¨Evolution of V Genes: DNA Sequence Structure of Functional Germ-Line Genes and Pseudogenes¨. Immunogenetics 42: 159-171. Sapp, J. (1987). Beyond the Genetic Cytoplasmic Inheritance and the Struggle for Authority in Genetics. Oxford University Press, New York, NY, USA. Sonneborn, T.M. (1930). Genetic studies on Stenostomum incaudatum. II. The effects of lead acetate on the hereditary constitution. J. Exp. Zool. 57: 409-439. Sonneborn, T.M. (1964). The differentiation of cells. Proc. Nat. Acad. Sci. USA 51: 915-929. Spadafora, C. (1998). Sperm cells and foreign DNA: a controversial relation. Bioessays 20: 955-964. Steele, E.J. (1979). Somatic Selection and Adaptive Evolution on the Inheritance of Acquired Characters (Williams and Wallace, eds.). 2nd edn. University of Chicago Press, Chicago, IL, USA. Steele, E.J. (Ed.) (1991). Somatic Hypermutation in V-regions. CRC Press, Boca Raton, FL, USA. Steele, E.J., Rothenfluh, H.S., Ada, G.L. and Blanden, R.V. (1993). Affinity maturation of lymphocyte receptors and positive selection of T cells in the thymus. Immunol. Rev. 135: 5-49. Steele, E.J., Blanden, R. and Rothenfluh, H. (1996). “How Have Antibody Genes Evolved?” Australas. Sci. 17: 46-49. Steele, E.J., Rothenfluh, H.S. and Blanden, R.V. (1997). Mechanism of antigen-driven somatic hypermutation of rearranged immunoglobulin V(D)J genes in the mouse. Immunol. Cell Biol. 75: 82-95. Steele, E.J., Lindley, R.A. and Blanden, R.V. (1998). Lamarck’s Signature, How Retrogenes are Changing Darwin’s Natural Selection Paradigm. Helix Books, Perseus Books, Reading, MA, USA. Steele, T. (1981). Lamarck and immunity; a conflict resolved. New Sci. 90: 360-361. Temin, H.M. (1989). Retrons in bacteria. Nature 339: 254-255. Waddington, C.H. (1953). Genetic assimilation of an acquired character. Evolution 7: 118-126. Waddington, C.H. (1957). The Strategy of the Genes. Allen and Unwin, London, England. Waddington, C.H. (1961). Genetic assimilation. Adv. Genet. 10: 257-293. Weiller, G.F., Rothenfluh, H.S., Zylstra, P., Gay, L.M., Averdunk, H., Steele, E.J. and Blanden, R.V. (1998). Recombination signature of germline immunoglobulin variable genes. Immunol. Cell Biol. 76: 179-185. Williams, G.C. (1996). Adaptation and Natural Selection. New Edition. Princeton University Press, Princeton, NJ, USA. Xiao, S., Zhang, Y. and Knoll, A.H. (1998). Three dimensional preservation of algae and animal embryos in a Neoproterozoicphosporite. Nature 391: 553-559. Zuckerkandl, E. (1986). Polite DNA: functional density and functional compatibility in genomes. J. Mol. Evol. 24: 12-27. Zuckerkandl, E. and Pauling, L. (1962). Molecular disease, evolution, and genic heterogeneity. In: Horizons in Biochemistry (Kasha, M. and Pullman, B., eds.). Academic Press, New York, NY, USA, pp. 189-225. |
|